

Page 1
R02/V2051
(1000522/1001210)
Page 2

2
Page 3

®
English
Meiosis, or reduction division, is a special type of cell division. Depending on the organism and cell type,
it can take anything from several days to years, resulting in the production of sex cells (gametes). Each
gamete precursor cell produces four gametes through reduction division. In general, there are two types
of gametes. Large, immobile cells are referred to as egg cells or oocytes, and small, mobile gametes as
sperm cells or spermatocytes.
Egg cells are formed by meiotic division from precursor cells in the ovaries. Human egg cells already begin
to mature in the embryo (3rd to 8th month of pregnancy), however, the cells remain in a meiotic intermediate phase until sexual maturity is reached. From then on, some of the immature egg cells complete the
meiotic division at regular intervals under the control of hormones. The maturation of human sperm cells
occurs regularly in the testes when sexual maturity has been reached. In this case, a complete meiotic
division takes 20-24 days.
Usually, body cells (e.g. precursor cells of the gametes) contain a double (diploid) chromosome complement, with one half of the chromosomes originating from the mother and the other half from the father.
Therefore, a twin copy exists of each chromosome, i.e. as a matching (homologous) pair of chromosomes.
By contrast, the gametes contain only a single (haploid) chromosome complement. In other words, egg or
sperm cells only contain half of the mother’s or father’s genetic information, so that when both cells join,
a new cell (zygote) with a complete diploid chromosome complement can originate.
The purpose of meiosis is therefore to reduce the normally diploid chromosome complement of a gamete
precursor cell to the haploid complement to establish the basis of any sexual reproduction. A further
important function of meiosis is to mix the genetic information. This is achieved by two mechanisms:
1. a random distribution of the maternal and paternal chromosomes to the sex cells being produced
2. the exchange of genes between the homologous chromosomes (genetic recombination)
(The underlying procedures of both mechanisms are explained below in the description of the individual
phases).
Meiosis
In humans, who have 23 chromosomes in the haploid complement, the random distribution of the
chromosomes alone allows for 2
of variations is furthermore increased by the exchange of genes between the chromosomes.
Prior to the meiotic division, the gamete precursor cells are in the interphase, which refers to the period
between two (mitotic or meiotic) cell divisions. The interphase comprises three stages:
• G1 phase (presynthesis) The stage where the cell grows.
• S phase (synthesis) In this phase, the centrioles and the DNA (deoxyribonucleic acid) begin to duplicate.
• G2 phase (postsynthesis) This phase separates the end of the DNA synthesis from the phase of division.
Furthermore, the duplication of the centrioles is completed.
Meiosis, the phase following the interphase, comprises two successive maturation (meiotic) divisions, which
are separated by a short, specific interphase (interkinesis). As in mitosis, several stages of division are differentiated in each meiotic division:
First meiotic division:
• Prophase I (four subsections: leptotene, zygotene, pachytene and diplotene with diakinesis)
• Metaphase I
• Anaphase I
• Telophase I
• Cytokinesis I
23
, i.e. 8.4 x 106 different genetic possibilities of variation. The number
3
Page 4

®
Meiosis
Interkinesis
Second meiotic division:
• Prophase II
• Metaphase II
• Anaphase II
• Telophase II
• Cytokinesis II
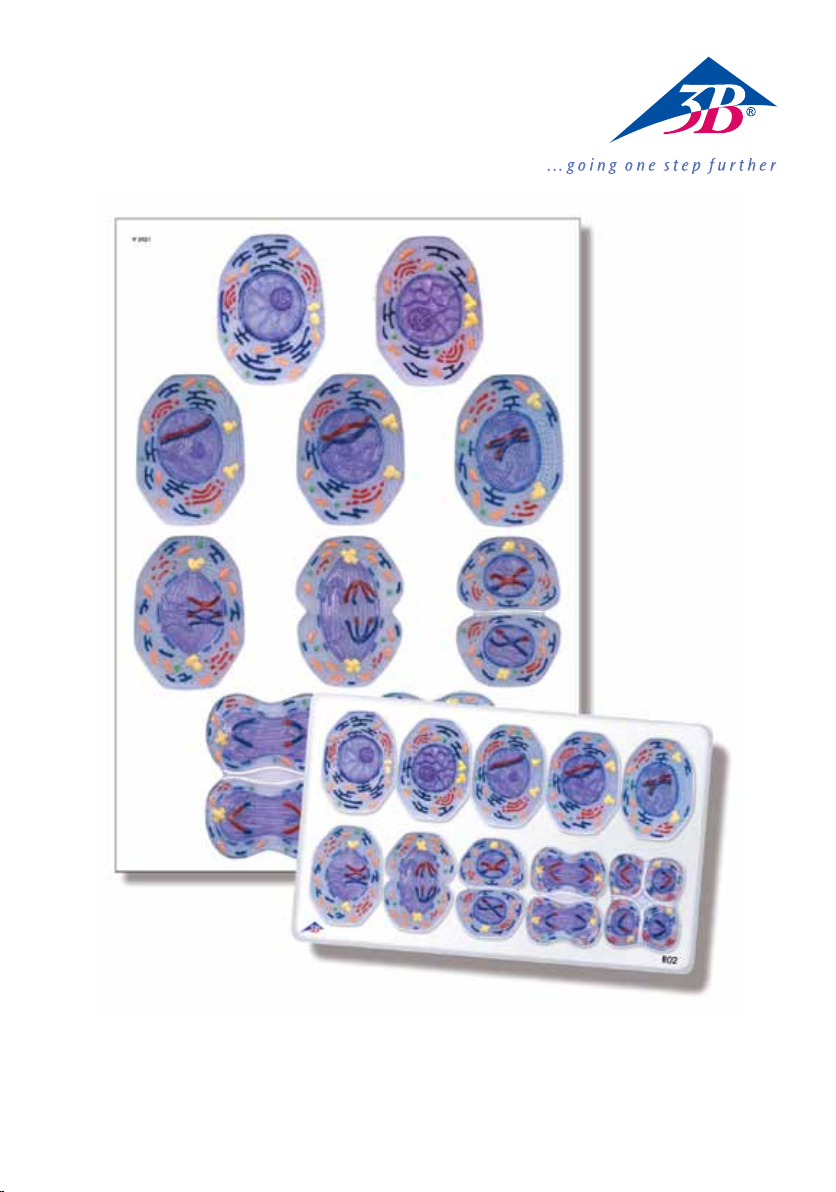
The 3B Scientific® model series on meiosis (product no. R02) and the wall chart on meiosis (V2051M,
V2051U) show a typical mammal cell at an enlargement of approx. 10,000 times. In the lower third
of the models/illustrations the cell organelles are portrayed open.
The 3B Scientific® model series on meiosis is supplied in a storage system, which is equipped with a
hanging device. The model series can thus be simply hung on a wall in order to save space. The models
also have magnets at the rear so that they can be arranged on magnetic boards in the classroom for teaching purposes.
At the end of this description you will find illustrations of the 10 phases included. You can use these to
make photocopies for your lessons. By colouring, labelling and correctly arranging the individual phases
your students can easily review and memorize what they have learned.
Free colour illustrations of the individual stages are also available on the Internet at www.3bscientific.com.
1. Interphase, stage of the G1 phase
Inside the cell the nucleus with the nucleolus (1) and its nuclear membrane (2) can be seen. The nucleus
also contains the not yet helical DNA (3) with the genetic information.
English
The cell itself receives its stability and shape from very fine tubes, the so-called microtubules (4) extending
through the cytoplasm. The microtubules control, among other things, the cell movements and the intracellular transport processes.
In the cytoplasm, the endoplasmic reticulum (5) can be seen. This is an intertwined tube system mainly in
charge of lipid synthesis, ion storage and redesigning and transporting certain proteins. The membrane of
the rough endoplasmic reticulum has ribosomes attached to it, which synthesize the proteins passing
through the endoplasmic reticulum.
The Golgi complex (or apparatus) (6) can also be referred to as “cell gland”. It is made up of stacks of
layered hollow sacs (Golgi cisternae), which swell up as the vesicles become too small and “pinch off”
(Golgi vesicles) (7). The Golgi complex receives membrane components and enzymes from the endoplasmic
reticulum. Its main function is collecting, packing and transporting secretions and producing lysosomes
(digestion vesicles) (8).
The main job of the lysosomes is breaking down cell components (= intracellular digestion).
The mitochondria (9) are in charge of producing energy for the cell.
The job of the centrioles (10) is to build up the cleavage spindle. They are hollow cylinders made up of
longitudinally arranged tubes (microtubules).
2. Prophase I
The prophase of the first meiotic division is the part of meiosis that takes longest. In the course of this
phase, chromosomes and chromatin change their structure and arrangement within the nucleus in a
4
Page 5

®
English
specific order. Therefore, prophase I is split up into four subsections (leptotene, zygotene, pachytene and
diplotene with diakinesis). In contrast to the mitotic prophase, which lasts several hours, meiotic prophase
I can take days, weeks, months or years.
Leptotene
At the beginning of prophase I (leptotene), the nucleolus (1) and the nuclear membrane (2) can be seen.
The chromosomes (3) are now visible as individual, long, thin threads. Its ends are attached within the
nuclear membrane. Each chromosome has already been replicated (duplicated) during the interphase and
is made up of two sister chromatids, which however are so close to each other that they cannot be differentiated. The centrioles were also duplicated in the interphase. Both pairs (4) begin to move in opposite
directions towards the two cell poles. Between them the so-called central spindle (5) begins to build up,
which consists of many microtubules.
3. Zygotene and Pachytene
One maternal (1) and one paternal homologue (2) (consisting of two sister chromatids) of one chromosome
pair are shown in different colours to represent the other chromosomes (2 x 23 in total).
Zygotene
The zygotene phase is initiated as soon as the homologous chromosomes begin to line up side by side to
form the synaptonemal complex (3) (parallel arrangement of the homologous partners). This process usually begins at the ends of the chromosomes and continues down to the other end, similar to a zipper. The
chromosome pairing (synapsis) occurs with high precision, so that the matching genes of the homologous
chromosomes face each other directly. This is an important requirement for the recombinant exchange of
gene sections (crossing over). The homologous chromosome pairs in meiotic prophase I are usually referred
to as bivalent, but since each homologous chromosome consists of the closely arranged sister chromatids,
they can also be referred to as tetrads.
Pachytene
As soon as all synaptonemal complexes are fully developed, i.e. the homologous chromosomes have all
lined up, the pachytene phase begins. From now on, recombination nodes (4) become visible at intervals
on the synaptonemal complexes, where the exchange of gene sections occurs.
Meiosis
4. Diplotene
After some gene sections have been exchanged, the homologous chromosomes (1) disjoin more and more,
remaining connected at one or more points of crossing over (chiasma bridges or chiasmata) (2). The chiasma bridges are the places where the genetic recombination (exchange of maternal and paternal genetic
information) occurred earlier. Egg cells can persist in the diplotene phase for months or even years.
5. Diakinesis
The end of meiotic prophase I begins when the chromosomes detach from the nuclear membrane (1). The
chromosomes are condensed and the sister chromatids, joined by centromeres (short DNA sequences with
a high AT level) (2), become visible. The non-sister-chromatids in which an exchange of gene sections occurred remain connected via chiasma bridges (3).
The phase following prophase I is metaphase I. The meiotic phases remaining at this point take up less
than 10% of the total time required for a complete meiosis.
6. Metaphase I
During the transition from prophase I into metaphase I, the centriole pairs (1) have reached the two oppo-
5
Page 6

®
Meiosis English
site cell poles. A spindle apparatus has developed and the nuclear membrane (2) dissolves. The chromosomes align at the equator level, forming the so-called metaphase plate. Viewed from the top, the chromosomes have a star-like shape (monaster or “mother“ star). The kinetochores (3) are protein complexes which
already have developed at the centromeres. A particularity of meiotic metaphase I is that the kinetochores
of each sister chromatid pair seem to have merged. The microtubules (4) of the central spindle, which now
have attached themselves precisely to the kinetochores of each sister chromatid pair (5), therefore all point
in the same direction. The chiasma bridges (6) are still intact. They play an important part in the correct
line-up of the homologous chromosomes at the equator level.
The endoplasmic reticulum (7) and the Golgi complex (8) have now been almost completely dissolved.
7. Anaphase I
In anaphase I of meiosis, the homologous chromosomes (1) disjoin and not, as in mitosis, the sister chromatids. In this process the chiasma bridges are dissolved, which so far held together the maternal and
paternal chromosomes.
Some mutant organisms, where meiotic crossing over occurs only on a limited level, have chromosome
pairs without chiasma bridges. These pairs are usually not fully disjoined (nondisjunction) and the resulting daughter cells have one chromosome too few or too many. Such malformations are referred to as
numerical chromosome aberrations, which cause deformities.
Disjunction begins at the kinetochores (2), the place where the traction fibres of the central spindle are
attached. From here, the chromosomes are pulled slowly towards the centrioles (4) located at the cell
poles, moving along the microtubules (3) which create a traction effect as they become shorter. The microtubules (5) that are not connected to chromosomes now become longer, thus increasing the distance between the centrioles and elongating the cell. At the equator level, the beginning stage of a cleavage furrow
(6) becomes visible.
The process of crossing over during the prophase and the random distribution of the maternal and paternal chromosomes to the cell poles result in a variation of the genetic information (ref. to introduction)
8. Telophase I, Cytokinesis I, Interkinesis, Prophase II and Metaphase II
Telophase I and Cytokinesis
In telophase I, the spindle disintegrates and a ring constriction (1) develops at the equator level. In addition, a thin nuclear membrane develops (2). During the following phase of cytokinesis, the cell body is divided exactly at the middle, at the ring constriction between the two new daughter nuclei (3). The daughter
nuclei each contain the maternal or paternal chromosome complement slightly varied through the process
of crossing over, with the DNA already present in duplicate, i.e. one chromosome consisting of two sister
chromatids (4).
The endoplasmic reticulum (5) and the Golgi complex (6) both have returned to their initial shape and size.
At the end of cytokinesis, the first meiotic division is completed.
Interkinesis
The first and second meiotic divisions are divided by a short resting period (interphase). However, there is
no duplication of the chromosomes consisting of two chromatids (no S phase). Both sister chromatids of
each chromosome remain connected by the centromeres (7).
6
Page 7

®
English
Meiotic division II
The second meiotic division occurs just like mitosis (usual nuclear and cell division). It is therefore also
referred to as equational division. Since the chromosomes were not duplicated again during the preceding
interkinesis, the second meiotic division, which now follows, includes the reduction of the genetic information to the haploid chromosome complement.
Prophase II
Prophase II is mostly like the prophase of mitosis and occurs very quickly in all organisms. The permeability
of the cell surface increases to allow the intake of surrounding liquids. The microtubule apparatus of the
cytoskeleton is reorganized. The nuclear membrane dissolves and the spindle is built up by rearranging
microtubules.
Metaphase II
In metaphase II, the chromosomes are once more arranged at the equator level and the two ends of the
spindle are located at the two opposite poles (as in metaphase I). A major difference to metaphase I is that
two kinetochores have developed at the sister chromatids which in this case point to opposite pole directions.
9. Anaphase II
During anaphase II, following now, the two sister chromatids (1) of each chromosome are disjoined just as
in mitosis.
The separation begins at the kinetochores (2), the point where the traction fibres of the central spindle
are attached. From here, the chromosomes are pulled slowly towards the centrioles (4) located at the cell
poles, moving along the microtubules (3) which create a traction effect as they become shorter. The microtubules (5) that are not connected to chromatids now become longer, thus increasing the distance between
the centrioles and elongating the cell. At the equator level, the beginning stage of a cleavage furrow (6)
becomes visible.
Meiosis
10. Telophase II and Cytokinesis II
The cleavage and division of the two cells produced during the first meiotic division now results in the production of four haploid cells (1) with different genetic combinations resulting from random chromosome
distribution and crossing over. This explains why siblings are not identical: one child has more features
from the father, the other more from the mother. It is also possible for features of the ancestors to reappear.
7
Page 8

®
DeutschMeiose
Die Meiose oder Reduktionsteilung ist eine besondere Form der Zellteilung. Abhängig vom Organismus und
Zelltyp kann sie einige Tage bis Jahre andauern und führt zur Bildung von Geschlechtszellen (Gameten). Aus
je einer Gametenvorläuferzelle entstehen durch Reduktionsteilung je vier Gameten. Es lassen sich generell
zwei Gametentypen unterscheiden. Große unbewegliche Zellen werden als Eizellen oder Oozyten bezeichnet und kleine bewegliche Gameten als Samenzellen oder Spermatozyten.
Eizellen entstehen durch meiotische Teilung aus Vorläuferzellen in den Eierstöcken (Ovarien). Die Reifung
der menschlichen Eizellen beginnt bereits im Embryo (3.-8. Schwangerschaftsmonat), wobei die Zellen
bis zur Geschlechtsreife in einem meiotischen Zwischenstadium verharren. Ab diesem Zeitpunkt vollenden
einige der unreifen Eizellen, in periodischen Abständen und unter Kontrolle von Hormonen, die meiotische
Teilung. Die Reifung menschlicher Samenzellen findet nach Einsetzen der Pubertät in regelmäßigen
Abständen in den Hoden (Testes) statt. Eine vollständige meiotische Teilung dauert in diesem Fall 20-24 Tage.
Normalerweise enthalten Körperzellen (z.B. Vorläuferzellen der Gameten) einen doppelten (diploiden)
Chromosomensatz, wobei eine Hälfte der Chromosomen von der Mutter und die andere Hälfte vom
Vater stammt. Jedes Chromosom liegt daher als Zwillingskopie vor, das heißt als sich entsprechendes
(homologes) Chromosomenpaar. Im Gegensatz dazu enthalten Gameten nur einen einfachen (haploiden)
Chromosomensatz. Anders ausgedrückt, Ei- oder Samenzellen enthalten nur die Hälfte des mütterlichen
bzw. väterlichen Erbgutes, so dass durch die Verschmelzung beider Zellen eine neue Zelle (Zygote) mit
komplettem, diploidem Chromosomensatz entstehen kann.
Der Sinn der Meiose besteht somit darin, den normalerweise diploiden Chromosomensatz einer Gametenvorläuferzelle auf den haploiden Satz zu reduzieren, um die Vorraussetzung für eine sexuelle Fortpflanzung
zu gewährleisten. Eine weitere wichtige Funktion der Meiose ist die Durchmischung des Erbgutes, welche
durch zwei Mechanismen erreicht wird: die zufällige Verteilung der mütterlichen und väterlichen Chromosomen auf die entstehenden Geschlechtszellen und den Austausch von Genen zwischen den homologen
Chromosomen (genetische Rekombination).
(Die zugrundeliegenden Prozesse beider Mechanismen werden später im Rahmen der Beschreibung der
einzelnen Stadien erklärt.)
Beim Menschen, mit einer Anzahl von 23 Chromosomen im haploiden Satz, ergeben sich allein durch die
zufällige Verteilung der Chromosomen 223 d.h. ca. 8,4 x 106 verschiedene genetische Variationsmöglichkeiten. Zudem wird die Variationsvielfalt durch den Austausch der Gene zwischen den Chromosomen
wesentlich erhöht.
Vor der meiotischen Teilung befinden sich die Gametenvorläuferzellen im Stadium der Interphase, die den
Zeitraum zwischen zwei Zellteilungen (mitotisch oder meiotisch) beschreibt. Die Interphase lässt sich in
drei Stadien gliedern:
• G1-Phase (Präsynthesephase) In diesem Stadium wächst die Zelle.
• S-Phase (Synthesephase) Hier beginnen sich die Centriolen und die DNS (Desoxyribonukleinsäure) zu
verdoppeln.
• G2-Phase (Postsynthesephase) Diese Phase trennt das Ende der DNS-Synthese von der Teilungsphase.
Zudem wird in diesem Stadium die Verdopplung der Centriolen abgeschlossen.
Die sich an die Interphase anschließende Meiose lässt sich in zwei aufeinanderfolgende Reifeteilungen
untergliedern, die durch eine kurze, spezielle Interphase (Interkinese) voneinander getrennt sind.
Wie bei der Mitose werden bei jeder Reifeteilung verschiedene Teilungsstadien unterschieden:
8
Page 9

®
Deutsch Meiose
Reifeteilung I
• Prophase I (vier Unterabschnitte: Leptotän, Zygotän, Pachytän und Diplotän mit Diakinese)
• Metaphase I
• Anaphase I
• Telophase I
• Zytokinese I
Interkinese
Reifeteilung II
• Prophase II
• Metaphase II
• Anaphase II
• Telophase II
• Zytokinese II
Die 3B Scientific® Modellserie zur Meiose (Produktnummer R02) bzw. in der Wandkarte zur Meiose
(V2051M, V2051U) zeigt eine typische Säugetierzelle in circa 10.000-facher Vergrößerung. Im unteren
Drittel der Modelle/Abbildungen sind die Zellorganellen eröffnet dargestellt.
Die 3B Scientific® Modellserie zur Meiose wird in einem Aufbewahrungssystem geliefert, das mit einer
Aufhängevorrichtung versehen ist. So können Sie die Modellserie auch einfach und platzsparend an einer
Wand aufhängen. Auf der Rückseite sind die Modelle mit Magneten versehen und können für den Unterricht an die Magnettafeln im Klassenzimmer angeordnet werden.
Am Ende dieser Beschreibung finden Sie Abbildungen der 10 dargestellten Stadien, die Sie als Kopiervorlage
für Ihren Unterricht nutzen können. Durch Ausmalen, Beschriften und richtiges Anordnen der einzelnen
Stadien können Ihre Schüler das Erlernte leicht nachvollziehen und vertiefen.
Farbige Abbildungen der einzelnen Stadien erhalten Sie auch kostenlos im Internet unter
www.3bscientific.com.
1. Interphase, Stadium der G1-Phase
Im Inneren der Zelle ist der Zellkern mit dem Nucleolus (Kernkörperchen) (1) und seiner Kernhülle (2) sichtbar. Im Zellkern befindet sich die noch entspiralisierte DNS (3) mit der genetischen Information.
Die Zelle selbst erhält ihre Stabilität und Form durch sehr dünne Röhren, die sogenannten Mikrotubuli (4),
die das Zellplasma durchspannen. Die Mikrotubuli steuern u.a. die Zellbewegung und die innerzellulären
Transportprozesse.
Im Zellplasma ist das endoplasmatische Retikulum (5) erkennbar. Dies ist ein verschlungenes Röhrensystem,
das vorwiegend der Lipidsynthese, als Ionenspeicher und der Umgestaltung und dem Transport bestimmter
Proteine dient. An der Membran des rauen endoplasmatischen Retikulums befinden sich Ribosomen, welche die Proteine synthetisieren, die durch das endoplasmatische Retikulum geschleust werden.
Den Golgi-Apparat (6) kann man auch als eine Art „Zelldrüse“ bezeichnen. Er besteht aus Stapeln von ineinander geschichteten Hohlkörpern (Golgi-Cisternen), die zu kleinen Bläschen auftreiben und sich abgliedern (Golgi-Vesikel) (7). Der Golgi-Apparat erhält vom endoplasmatischen Retikulum Membranbausteine
und Enzyme angeliefert. Seine Hauptaufgabe besteht im Sammeln, Verpacken und Abtransport von Sekreten
und in der Bildung von Lysosomen (Verdauungsbläschen) (8). Hauptaufgabe der Lysosomen ist der Abbau
von Zellbestandteilen. (=intrazelluläre Verdauung). Für die Energiegewinnung der Zelle sind die Mitochondrien (9) zuständig. Aufgabe der Centriolen (10) ist es, die Teilungsspindel aufzubauen. Sie sind Hohlzylinder,
die aus längsverlaufenden Röhren (Mikrotubuli) gebildet werden.
9
Page 10

®
Meiose Deutsch
2. Prophase I
Die Prophase der ersten Reifeteilung stellt den zeitlich längsten Abschnitt der Meiose dar. Im Verlauf
dieser Phase verändern Chromosomen und Chromatin in einer bestimmten Reihenfolge ihre Struktur und
Anordnung im Zellkern. Daher wird die Prophase I in vier Unterabschnitte (Leptotän, Zygotän, Pachytän
und Diplotän mit Diakinese) gegliedert. Im Gegensatz zur mitotischen Prophase, welche einige Stunden
andauert, kann die meiotische Prophase I Tage, Wochen, Monate oder Jahre beanspruchen.
Leptotän
Zu Beginn der Prophase I (Leptotän) sind der Nucleolus (Zellkörperchen) (1) und die Kernhülle (2) zu erkennen. Die Chromosomen (3) werden nun als einzelne, lange, dünne Fäden sichtbar. Ihre Enden sind in der
Kernmembran verankert. Jedes Chromosom wurde bereits in der Interphase repliziert (verdoppelt) und
besteht aus zwei Schwester-Chromatiden, die allerdings so eng beieinander liegen, dass sie nicht voneinander zu unterscheiden sind. Die Centriolen wurden ebenfalls in der Interphase verdoppelt. Beide Paare (4)
beginnen voneinander weg in Richtung der beiden Zellpole zu wandern. Zwischen ihnen beginnt sich die
sogenannte Zentralspindel (5) auszubilden, die aus vielen Mikrotubuli besteht.
3. Zygotän und Pachytän
Je ein mütterliches (1) und ein väterliches Homolog (2) (bestehend aus zwei Schwester-Chromatiden) eines
Chromosomenpaares, sind in unterschiedlichen Farben stellvertretend für die anderen Chromosomen (insgesamt 2 x 23) dargestellt.
Zygotän
Die Phase des Zygotäns wird eingeleitet sobald die homologen Chromosomen damit anfangen, sich dicht
aneinander zu lagern, um den synaptonemalen Komplex (3) (Parallelanordnung der homologen Partner)
auszubilden. Meist beginnt dieser Prozess an den Enden der Chromosomen und wird, ähnlich wie bei
einem Reißverschluss, zum anderen Ende hin fortgesetzt. Die Chromosomenpaarung (Synapsis) erfolgt mit
hoher Präzision, so dass die sich entsprechenden Gene der homologen Chromosomen direkt gegenüberliegen. Dieses ist eine wichtige Voraussetzung für den rekombinanten Austausch der Genabschnitte (Crossing
over). Jedes homologe Chromosomenpaar in der meiotischen Prophase I wird für gewöhnlich Bivalent
genannt, da aber jedes einzelne homologe Chromosom aus den eng beieinanderliegenden SchwesterChromatiden besteht, kann man auch von Tetraden sprechen.
Pachytän
Sobald alle synaptonemalen Komplexe voll ausgebildet sind, d.h. sich die homologen Chromosomen
vollständig aneinandergelagert haben, ist die Phase des Pachytäns erreicht. Auf den synaptonemalen
Komplexen werden nun in Abständen Rekombinationsknoten (4) sichtbar, in denen der Austausch von
Genabschnitten erfolgt.
4. Diplotän
Nach dem Austausch einiger Genabschnitte lösen sich die homologen Chromosomen (1) zunehmend voneinander aber bleiben zunächst an einem oder mehreren Überkreuzungspunkten verbunden (Chiasmabrücken oder Chiasmata) (2). An den Chiasmabrücken hat zuvor die genetische Rekombination (Austausch
von mütterlicher und väterlicher Erbinformation) stattgefunden. Eizellen können monate- bis jahrelang
im Zustand des Diplotäns verharren.
5. Diakinese
Das Ende der meiotischen Prophase I wird durch das Ablösen der Chromosomen von der Kernhülle (1) eingeleitet. Die Chromosomen verdichten sich und die Schwester-Chromatiden, die über Centromere (kurze,
AT-reiche Sequenzen der DNS) (2) verbunden sind, werden sichtbar. Die Nicht-Schwester-Chromatiden, in
10
Page 11

®
Deutsch Meiose
denen ein Austausch von Genabschnitten stattgefunden hat, sind auch weiterhin über Chiasmabrücken (3)
miteinander verbunden.
Die Prohase I geht in die Metaphase I über. Die nun noch verbleibenden meiotischen Phasen beanspruchen
weniger als 10 % der erforderlichen Gesamtzeit einer kompletten Meiose.
6. Metaphase I
Am Übergang der Prophase I in die Metaphase I sind die Centriolenpaare (1) an den beiden gegenüberliegenden Polen der Zelle angekommen. Ein Spindelapparat hat sich ausgebildet und die Kernhülle (2) zerfällt. Die Chromosomen ordnen sich in der Äquatorialebene an. Sie bilden die sogenannte Metaphasenplatte. In der Aufsicht erscheinen die Chromosomen als sternförmige Gebilde (Monaster oder Mutterstern).
Die Kinetochoren (3) sind Proteinkomplexe, welche sich bereits an den Centromeren ausgebildet haben.
Eine Besonderheit der meiotischen Metaphase I ist, dass die Kinetochoren eines jeden SchwesterChromatidenpaares fusioniert zu sein scheinen. Die Mikrotubuli (4) der Zentralspindel, die jetzt exakt an
den Kinetochoren eines jeden Schwester-Chromatidenpaares (5) angesetzt haben, weisen daher alle in die
gleiche Richtung. Die Chiasmabrücken (6) sind immer noch erhalten. Sie spielen eine wichtige Rolle für das
korrekte Aufreihen der homologen Chromosomen in der Äquatorialebene.
Das endoplasmatische Retikulum (7) und der Golgi-Apparat (8) sind jetzt fast vollständig aufgelöst.
7. Anaphase I
In der Anaphase I der Meiose trennen sich die homologen Chromosomen (1) und nicht wie in der Mitose
die Schwester-Chromatiden voneinander. Hierbei werden die Chiasmabrücken, die die homologen mütterlichen und väterlichen Chromosomen zusammenhielten, aufgelöst.
Einige mutante Organismen, bei denen das meiotische Crossing over nur in begrenztem Maße auftritt, weisen Chromosomenpaare ohne Chiasmabrücken auf. Diese Paare werden meistens nicht richtig voneinander
getrennt (nondisjunction) und die resultierenden Tochterzellen enthalten ein Chromosom zu wenig, bzw.
zuviel. Man bezeichnet eine solche Fehlbildung als numerische Chromosomenaberration, die zu
Fehlbildungen führt.
Die Trennung beginnt an den Kinetochoren (2), der Anheftungsstelle der Zugfasern der Zentralspindel.
Von dort aus werden die Chromosomen dann langsam über die sich verkürzenden Mikrotubuli (3) und
die dadurch entstehende Zugwirkung zu den an den Zellpolen liegenden Centriolen (4) gezogen. Die Mikrotubuli (5), die nicht mit Chromosomen verbunden sind, werden jetzt länger, wodurch sich der Abstand zwischen den Centriolen vergrößert und die Zelle in die Länge gezogen wird. In der Äquatorialebene ist die
Andeutung einer Schnürfurche (6) erkennbar.
Durch das in der Prophase erfolgte Crossing over und die zufällige Verteilung der mütterlichen und väterlichen Chromosomen zu jeweils einem Zellpol, kommt es zu einer Variation des Erbgutes (siehe Einleitung).
8. Telophase I, Zytokinese I, Interkinese, Prophase II und Metaphase II
Telophase I und Zytokinese
In der Telophase I bildet sich der Spindelapparat zurück und ein Schnürring (1) entsteht in der Äquatorialebene. Ebenso bildet sich eine dünne Kernmembran (2). Der Zellleib wird während der sich anschließenden Zytokinese genau in der Mitte, am Schnürring zwischen den beiden neu entstandenen Tochterkernen
(3), durchtrennt. Die Tochterkerne enthalten jeweils den durch das Crossing over leicht veränderten mütterlichen bzw. väterlichen Chromosomensatz, wobei die DNS bereits verdoppelt vorliegt, d.h. ein Chromosom besteht aus zwei Schwester-Chromatiden (4).
11
Page 12

®
DeutschMeiose
Das endoplasmatische Retikulum (5) und der Golgi-Apparat (6) haben ihre anfängliche Form und Größe
wiedererlangt.
Am Ende der Zytokinese ist die erste Reifeteilung beendet.
Interkinese
Die erste und zweite Reifeteilung sind durch eine kurze Ruhepause (Interphase) voneinander getrennt. Eine
Verdopplung der aus zwei Chromatiden bestehenden Chromosomen erfolgt allerdings nicht (keine S-Phase).
Durch die Centromere (7) bleiben die beiden Schwester-Chromatiden jedes einzelnen Chromosoms miteinander verbunden.
Reifeteilung II
Die zweite Reifeteilung verläuft wie eine Mitose (übliche Kern- und Zellteilung). Deswegen wird sie auch
Äquationsteilung genannt. Da die Chromosomen während der vorangegangenen Interkinese nicht wieder
verdoppelt wurden, kommt es bei der nun folgenden zweiten meiotischen Teilung zur Reduktion des
Erbgutes auf den haploiden Chromosomensatz.
Prophase II
Die Prophase II entspricht weitgehend der Prophase einer Mitose und verläuft in allen Organismen sehr
schnell. Die Durchlässigkeit der Zelloberfläche wird erhöht, um Flüssigkeit aus der Umgebung aufzunehmen.
Der Mikrotubulusapparat des Zytoskeletts ordnet sich um. Die Kernmembran wird aufgelöst und die Spindel
bildet sich aus sich neu organisierenden Mikrotubuli.
Metaphase II
In der Metaphase II sind die Chromosomen wiederum in der Äquatorialebene angeordnet und die beiden
Spindelenden liegen an den gegenüberliegenden Polen (analog zur Metaphase I). Ein wesentlicher Unterschied zu Metaphase I besteht darin, dass sich an den Schwester-Chromatiden zwei Kinetochoren entwickelt haben, die diesmal in gegensätzliche Polrichtungen weisen.
9. Anaphase II
In der nun folgenden Anaphase II werden die beiden Schwester-Chromatiden (1) jedes Chromosoms wie in
der Mitose voneinander getrennt.
Die Trennung beginnt an den Kinetochoren (2), der Anheftungsstelle der Zugfasern der Zentralspindel.
Von dort aus werden die Chromatiden dann langsam über die sich verkürzenden Mikrotubuli (3) und die
dadurch entstehende Zugwirkung zu den an den Zellpolen liegenden Centriolen (4) gezogen. Die Mikrotubuli (5), die nicht mit Chromatiden verbunden sind, werden jetzt länger, wodurch sich der Abstand zwischen den Centriolen vergrößert und die Zelle in die Länge gezogen wird. In der Äquatorialebene ist die
Andeutung einer Schnürfurche (6) erkennbar.
10. Telophase II und Zytokinese II
Durch die Abschnürung und Teilung der beiden aus der ersten Reifeteilung hervorgegangenen Zellen,
entstehen jetzt vier haploide Zellen (1) mit unterschiedlicher, durch zufällige Chromosomenaufteilung
und Crossing over verursachter Genkombination. Hieraus erklärt sich die Beobachtung, dass Geschwister
nicht identisch sind: das eine Kind lässt mehr die Merkmale des Vaters, das andere Kind die Merkmale
der Mutter erkennen. Es können dadurch auch wieder Merkmale der Vorfahren auftauchen.
12
Page 13

®
Español Meiosis
La meiosis o división reduccional es una forma de división celular especial. En función del organismo y
tipo de célula puede durar entre varios días y varios años y resulta la formación de células sexuales
(gametos). De una célula precursora de gametos se forman cuatro gametos, respectivamente, por división
reduccional. Generalmente, se distinguen dos tipos de gametos. Las células grandes inmóviles se denominan óvulos u oocitos y los gametos pequeños y móviles se denominan espermatozoides o espermatocitos.
Los óvulos se forman por división meiótica a partir de células precursoras en los ovarios. La maduración de
los óvulos humanos ya empieza en el embrión (3.-8. mes de embarazo), permaneciendo las células en un
estado intermedio meiótico hasta la pubertad. A partir de este momento, algunas de las células inmaduras
acaban la división meiótica, a intervalos periódicos y bajo el control de hormonas. La maduración de los
espermatozoides humanos comienza en la pubertad y se produce a intervalos regulares en los testículos.
En este caso, una división meiótica completa dura 20-24 días.
Normalmente, las células corporales (p. ej. células precursoras de gametos) contienen un juego cromosómico doble (diploide), procediendo la mitad de los cromosomas de la madre y la otra mitad del padre. Por
lo tanto, cada cromosoma existe como copia gemela, es decir como par de cromosomas que se corresponden (homólogos). Al contrario, los gametos contienen un solo juego cromosómico simple (haploide). O por
decirlo así, los óvulos o espermatozoides contienen solamente la mitad de la herencia de la madre y del
padre, respectivamente, de manera que por la fusión de ambas células se pueda formar una nueva célula
(cigoto) con un juego cromosómico diploide completo.
Por lo tanto, la meiosis tiene la finalidad de reducir el juego cromosómico de una célula precursora de
gametos que normalmente es diploide a un juego haploide para garantizar el requisito de la reproducción sexual. Otra función importante de la meiosis es entremezclar la herencia, lo que se consigue por dos
mecanismos:
1. la distribución al azar de los cromosomas maternos y paternos entre las células sexuales en vías de
formación
2. el intercambio de genes entre los cromosomas homólogos (recombinación genética)
(Los procesos en los cuales se basan ambos mecanismos se explicarán más abajo, en el marco de la descripción de las diferentes fases.)
En el ser humano, con un número de 23 cromosomas en el juego haploide, resultan ya sólo por la distribución al azar de los cromosomas unas 223, o sea 8,4 x 106 diferentes variaciones genéticas posibles. Además,
la diversidad de variaciones posibles es aumentada considerablemente por el intercambio de genes entre
los cromosomas.
Antes de la división meiótica, las células precursoras de gametos se encuentran en el estado de la interfase,
que describe el período comprendido entre dos divisiones celulares (mitóticas o meióticas). La interfase
comprende los períodos siguientes:
• Fase G1 (fase presintética) Durante esta fase la célula crece.
• Fase S (fase sintética) Aquí los centríolos y el ADN (ácido desoxirribonucleico) empiezan a duplicarse.
• Fase G2 (fase postsintética) Esta fase separa el final de la síntesis del ADN de la fase de división.
Además, en esta fase se concluye la duplicación de los centríolos.
La meiosis que sigue inmediatamente a la interfase consta de dos divisiones meióticas sucesivas intercaladas por una corta interfase específica (intercinesis). Igual que en la mitosis, cada división meiótica consta
de distintos períodos de división:
13
Page 14

®
EspañolMeiosis
División meiótica I
• Profase I (cuatro subdivisiones: leptoteno, anfiteno, paquiteno y diploteno con diacinesis)
• Metafase I
• Anafase I
• Telofase I
• Citocinesis I
Intercinesis
División meiótica II
• Profase II
• Metafase II
• Anafase II
• Telofase II
• Citocinesis II
En la serie de modelos relativos a la meiosis de 3B Scientific® (N° de producto R02) o el mapa mural relativo
a la meiosis (V2051M, V2051U), respectivamente, se representa una celúla típica de un mamífero a 10.000
aumentos aprox.. En el tercio inferior de los modelos/figuras se representan de forma abierta las organelas
celulares.
La serie de modelos de 3B Scientific® relativa a la meiosis se suministra en el interior de un sistema para
guardarlos que está provisto de un dispositivo para colgarlo. Así Vd. podrá asimismo colgar fácilmente la
serie de modelos en un muro, lo que ahorra espacio. Los modelos llevan imán en el dorso y por lo tanto,
en clase podrán colocarse en los tablones magnéticos de las aulas.
Al final de la presente descripción Vd. encontrará unas figuras de las 10 fases representadas que podrá
emplear como patrones para sacar copias para sus clases. Al pintar y poner una leyenda en la respectiva
fase y colocarlas correctamente, sus alumnos podrán fácilmente entender y profundizar en lo aprendido.
Las figuras en color de las respectivas fases también son disponibles gratuitamente en Internet, en
www.3bscientific.com.
1. Interfase, período de la fase G
En el interior de la célula se observa el núcleo celular con el nucléolo (1) y su membrana nuclear (2). En el
interior del núcleo celular se halla el ADN que aún no tiene forma de hélice (3) con la información genética.
La célula conserva su estabilidad y forma mediante tubos muy delgados, los llamados microtúbulos (4) que
atraviesan el citoplasma. Los microtúbulos controlan el movimiento celular y los procesos de transporte
intracelulares, entre otros.
En el interior del citoplasma se observa el retículo endoplasmático (5). Es un sistema tubular enredado que
sirve sobre todo para la síntesis de lípidos, como almacén de iones y para la transformación y el transporte
de determinadas proteínas. En la membrana del retículo endoplasmático rugoso se hallan ribosomas que
sintetizan las proteínas que pasan por el retículo endoplasmático.
El aparato de Golgi (6) puede denominarse asimismo una especie de “glándula celular”. Consta de pilas de
cuerpos huecos entremetidos (cisternas de Golgi) que se empujan hacia arriba, formando pequeñas vesículas que posteriormente se separan (vesículas de Golgi) (7). El aparato de Golgi recibe del retículo endoplasmático materiales para la membrana y enzimas que éste le suministra. Su tarea principal es la de recoger,
empaquetar y transportar secreciones y fabricar lisosomas (vesículas de digestión) (8).
La tarea principal de los lisosomas es la degradación de componentes celulares (=digestión intracelular).
Los responsables de la producción de energía en la célula son las mitocondrias (9).
La tarea de los centríolos (10) es la formación del huso acromático. Son cilíndros huecos formados por
tubos longitudinales (microtúbulos).
14
1
Page 15

®
Español
2. Profase I
La profase de la primera división meiótica es la fase más larga de la meiosis. Durante esta fase, los
cromo-somas y la cromatina cambian, en un orden determinado, de estructura y colocación en el interior
del núcleo celular. Por eso, la profase I se subdivide en cuatro períodos (leptoteno, anfiteno, paquiteno y
diploteno con diacinesis). Contrariamente a la profase mitótica que dura varias horas, la profase meiótica I
puede durar varios días, semanas, meses o años.
Leptoteno
Al principio de la profase I (leptoteno) se observan el nucléolo (1) y la membrana nuclear (2). Ahora los
cromosomas (3) se hacen visibles como largas hebras delgadas sueltas. Sus extremidades son asociadas a la
membrana nuclear. Cada cromosoma ya ha sido replicado (duplicado) durante la interfase y está conformado
por dos cromátides hermanas, que, sin embargo, están tan cerca uno del otro que no se pueden distinguir.
Los centríolos también han sido duplicados durante la interfase. Ambos pares (4) empiezan a alejarse uno
del otro, trasladándose hacia los polos opuestos de la célula. Entre ellos, comienza a formarse el llamado
huso central (5) que está formado por numerosos microtúbulos.
3. Anfiteno y Paquiteno
Un homólogo materno (1) y un homólogo paterno (2) (constituidos por dos cromátides hermanas) de un
par de cromosomas, respectivamente, son representativos de los demás cromosomas (en total 2 x 23) y
representados en distintos colores.
Anfiteno
Se inicia la fase del anfiteno, cuando los cromosomas homólogos comiencen a aparearse estrechamente
para formar el complejo sinaptonémico (3) (colocación paralela de los asociados homólogos). En la mayoría
de los casos, este proceso comienza en las extremidades de los cromosomas y continúa en dirección a la
extremidad opuesta, de forma parecida al funcionamiento de una cremallera. El apareamiento de cromosomas (sinapsis) se realiza con alta precisión, de manera que los genes de los cromosomas homólogos
que se correspondan estén situados exactamente uno en frente de otro. Esto es un requisito importante
del intercambio recombinante de los segmentos cromosómicos (crossing-over). A cada par de cromosomas homólogos de la profase meiótica I se suele denominar bivalente, pero dado que cada cromosoma
homólogo individual está conformado por las cromátides hermanas situadas muy cerca una de otra, también pueden llamarse tetradas.
Meiosis
Paquiteno
Cuando todos los complejos sinaptonémicos se hayan completamente formados, es decir cuando los cromosomas homólogos se hayan completamente unidos, ha llegado la fase del paquiteno. Entonces, sobre
los complejos sinaptonémicos se hacen visibles por intervalos nudos de recombinación (4), en los cuales
se realiza el intercambio de segmentos cromosómicos.
4. Diploteno
Después del intercambio de varios segmentos cromosómicos, los cromosomas homólogos (1) se separan
progresivamente, aunque por de pronto quedan unidos por uno o varias puntos de entrecruzamiento
(puentes de quiasma o quiasmas) (2). En los quiasmas se realizó previamente la recombinación genética
(intercambio de información genética materna y paterna). Los óvulos pueden permanecer durante meses
o años en el estado del diploteno.
5. Diacinesis
El final de la profase meiótica I es introducida por la separación de los cromosomas de la membrana
nuclear (1). Los cromosomas se condensan y las cromátides hermanas que están unidos por centrómeros
(secuencias cortas del ADN, ricas en adenina y timina) (2) se hacen visibles. Las cromátides no hermanas,
en las cuales se efectuó un intercambio de segmentos cromosómicos, quedan unidas por quiasmas (3).
15
Page 16

®
EspañolMeiosis
De la profase I se pasa a la metafase I. Las restantes fases meióticas requieren menos del 10% del tiempo
total requerido para la meiosis completa.
6. Metafase I
En el momento de la transición de la profase I a la metafase I, los pares de centríolos (1) han llegado a los
dos polos opuestos de la célula. Se ha formado el huso acromático y la membrana nuclear (2) se descompone. Los cromosomas se disponen en el plano ecuatorial. Forman el llamado plato de metafase. A vista
desde arriba, los cromosomas son parecidos a estrellas (monáster o estrella madre). Los cinetocoros (3) son
complejos proteínicos que ya se han formado en los centrómeros. Una particularidad de la metafase meiótica I es que los cinetocoros de cada par de cromátides hermanas parecen estar fusionados. Por consiguiente, los microtúbulos (4) del huso central que entonces se han establecido exactamente en los cinetocoros
de cada par de cromátides hermanas (5), se dirigen todos en la misma dirección. Los quiasmas (6) siguen
existiendo. Desempeñan un papel importante en la alineación correcta de los cromosomas homólogos en
el plano ecuatorial.
Ahora el retículo endoplasmático (7) y el aparato de Golgi (8) están casi completamente descompuestos.
7. Anafase I
Durante la anafase I de la meiosis, se separan los cromosomas homólogos (1) y no las cromátides hermanas
como se produce en la mitosis. Durante este proceso se desintegran los quiasmas que unían los cromosomas homólogos maternos y paternos.
Algunos organismos mutantes, en los que el crossing-over meiótico no se produce que en medida limitada,
presentan pares de cromosomas sin quiasmas. En la mayoría de los casos, estos pares no se separan correctamente (nondisjunction) y las células hijas resultantes tienen un cromosoma de menos o de más, respectivamente. Tal deformación se denomina aberración cromosomática numérica que conlleva deformaciones.
La separación comienza en los cinetocoros (2), punto donde se adhieren las fibras de tracción del huso
central. Desde allá, los cromosomas son tirados paulatinamente a través de los microtúbulos (3) que se
contraen y la consiguiente fuerza de tracción hacia los centríolos (4) situados en los polos celulares. Los
microtúbulos (5) que no están unidos a cromosomas, se hacen más largos ahora, por lo cual aumenta la
distancia entre los centríolos y se alarga la célula. En el plano ecuatorial se observa un somero surco de
constricción (6).
El crossing-over realizado durante la profase y la distribución al azar de los cromosomas maternos y paternos
con destino a uno de los polos celulares, respectivamente, conlleva una variación de la herencia (véase
Introducción).
8. Telofase I, Citocinesis I, Intercinesis, Profase II y Metafase II
Telofase I y Citocinesis
Durante la telofase I se desorganiza el huso acromático y se forma un anillo de constriccion (1) en el plano
ecuatorial. Asimismo se forma una membrana nuclear delgada (2). Durante la citocinesis subsi-guiente, el
cuerpo de la célula se divide en dos exactamente por el medio, en el anillo de constricción, entre los dos
núcleos de las células hijas recién formados (3). Los núcleos de las células hijas contienen el juego de cromosomas materno o paterno, respectivamente, modificados levemente por el crossing-over, existiendo el
ADN ya por duplicado, es decir un cromosoma está formado por dos cromátides hermanas (4).
El retículo endoplasmático (5) y el aparato de Golgi (6) han recuperado su forma y tamaño iniciales.
Al final de la citocinesis, la primera división meiótica está terminada.
16
Page 17

®
Español Meiosis
Intercinesis
La primera y la segunda división meiótica son intercaladas por un breve descanso (interfase). Sin embargo,
no se realiza ninguna duplicación de los cromosomas formados por dos cromátides (no hay fase S). Por los
centrómeros (7), ambas cromátides hermanas de cada cromosoma quedan unidas entre sí.
División meiótica II
La segunda división meiótica se produce de la misma manera que la mitosis (división nuclear y celular
normal). Por este motivo, se denomina asimismo división ecuacional. Como los cromosomas no han sido
duplicados durante la intercinesis que precede, resulta la reducción de la herencia al juego de cromosomas
haploide durante la segunda división meiótica.
Profase II
La profase II equivale en gran parte a la profase de la mitosis y se produce muy rápidamente en cualquier
organismo. La permeabilidad de la superficie celular aumenta para poder absorber la humanidad del
ambiente. El aparato microtubular del citoesqueleto se reorganiza. La membrana nuclear se desintegra y
se forma el huso a partir de microtúbulos que se están reorganizando.
Metafase II
Durante la metafase II, los cromosomas vuelven a disponerse en el plano ecuatorial y las dos extremidades
del huso se hallan en los polos opuestos (análogamente a la metafase I). Sin embargo, difiere considerablemente de la metafase I por el hecho de que en las cromátides hermanas se han formado dos cinetocoros
que ésta vez se dirigen en unas direcciones polares opuestas.
9. Anafase II
A continuación, durante el anafase II, las dos cromátides hermanas (1) de cada cromosoma se separan,
igual que en la mitosis.
La separación comienza en los cinetocoros (2), punto donde se adhieren las fibras de tracción del huso central. Desde allá, los cromátides son tirados paulatinamente a través de los microtúbulos (3) que se contraen
y la consiguiente fuerza de tracción hacia los centríolos (4) situados en los polos celulares. Los microtúbulos (5) que no están unidos a cromátides, se hacen más largos ahora, por lo cual aumenta la distancia entre
los centríolos y se alarga la célula. En el plano ecuatorial se observa un somero surco de constricción (6).
10. Telofase II y Citocinesis II
De la constricción y la división de las dos células procedentes de la primera división meiótica, resultan
ahora cuatro células haploides (1) con distintas combinaciones genéticas debidas a la distribución al azar
de los cromosomas y el crossing-over. Eso explica el hecho observado que los hermanos no son idénticos:
en el uno se observan más las características del padre, en el otro las de la madre. Por este motivo, también es posible que resurgan características de los antepasados.
17
Page 18

®
Français
La méiose ou la division réductrice est une forme particulière de la division cellulaire. En fonction de
l’organisme et du type de cellule, sa durée est de quelques jours à quelques années, et elle entraîne la formation de cellules sexuées (gamètes). De chaque cellule précurseur d’un gamète résultent quatre gamètes
par la division réductrice. En général, on distingue deux types de gamètes. Les grandes cellules immobiles
sont désignées par le terme d’ovules ou d’ovocytes et les petits gamètes mobiles par le terme de spermatozoïdes ou spermatocytes.
Les ovules résultent de la division méiotique des cellules précurseurs qui a lieu dans les ovaires.
La maturation des ovules humains débute déjà dans l’embryon (du 3ème au 8ème mois de la grossesse),
les cellules stagnent à un stade méiotique intermédiaire jusqu’à la puberté. A partir de ce moment, certains des ovules immatures accomplissent la division méiotique à intervalles réguliers et sous le contrôle
des hormones. La maturation des spermatozoïdes humains a lieu à intervalles réguliers dans les testicules
après la survenue de la puberté. Une division méiotique complète dure dans ce cas 20 à 24 jours.
Normalement, les cellules corporelles (p.ex. les cellules précurseurs des gamètes) contiennent un double
jeu de chromosomes (diploïde) dont une moitié des chromosomes provient de la mère et l’autre moitié du
père. Chaque chromosome est par conséquent présent sous forme de copie jumelle, c’est-à-dire en tant
que paire (homologue) de chromosomes correspondants. Les gamètes contiennent au contraire uniquement un simple jeu de chromosomes (haploïde). En d’autres termes, les ovules ou les spermatozoïdes contiennent seulement la moitié du génotype maternel et paternel, tant et si bien que de la fusion des deux
cellules peut résulter une nouvelle cellule (zygote) possédant un jeu de chromosomes diploïde complet.
La fonction de la méiose consiste par conséquent à réduire le jeu de chromosomes normalement diploïde
d’une cellule précurseur d’un gamète à un jeu haploïde afin d’assurer la condition pour la reproduction
sexuelle. Une autre fonction importante de la méiose réside dans le mélange du génotype, réalisé par deux
mécanismes :
1. la répartition fortuite des chromosomes maternels et paternels sur les cellules sexuées en formation
2. l’échange des gènes entre les chromosomes homologues (recombinaison génétique)
(Les processus de base de ces deux mécanismes sont expliqués plus loin dans le cadre de la description de
chaque stade).
Chez l’homme, avec un nombre de 23 chromosomes dans le jeu haploïde, la répartition fortuite des chromosomes entraîne 223, c.-à-d. env. 8,4 x 106 possibilités différentes de variations génétiques. Par ailleurs,
la multitude de variations augmente considérablement par l'échange des gènes entre les chromosomes.
Avant la division méiotique, les cellules précurseurs des gamètes se trouvent au stade de l’interphase qui
décrit la période entre deux divisions cellulaires (mitotique ou méiotique). L’interphase se divise en trois
stades :
• Phase G1 (phase de présynthèse) A ce stade, la cellule croît.
• Phase S (phase de synthèse) C’est durant cette phase que les centrioles et l’ADN
(acide désoxyribonucléique) commencent à se dédoubler.
• Phase G
La méiose succédant à l’interphase se divise en deux divisions de maturation successives, séparées par une
interphase spéciale et courte (intercinèse). De même que lors de la mitose, on distingue différents stades
de division lors de chaque division de maturation :
18
(phase de postsynthèse) Cette phase sépare la fin de la synthèse de l’ADN de la phase de
2
division. Par ailleurs, le dédoublement des centrioles est achevé durant ce stade.
Page 19

®
Français Méiose
Division de maturation I
• Prophase I (quatre sous-sections : leptotène, zygotène, pachytène et diplotène avec diacinèse)
• Métaphase I
• Anaphase I
• Télophase I
• Cytocinèse I
Intercinèse
Division de maturation II
• Prophase II
• Métaphase II
• Anaphase II
• Télophase II
• Cytocinèse II
La série des modèles 3B Scientific® de la méiose (numéro de produit R02) ainsi que la table murale
“La méiose” (V2051M, V2051U) montrent une cellule typique des mammifères, agrandie environ 10.000
fois. Dans le tiers inférieur des modèles/représentations, les organelles cellulaires ouverts sont représentés.
La série des modèles 3B Scientific® de la méiose est fournie dans un système de conservation muni d’un
dispositif de suspension, ce qui vous permet de suspendre simplement la série des modèles au mur en
économisant de la place. Au verso, les modèles sont pourvus d’aimants, ce qui permet de les classer sur
les tableaux magnétiques dans la classe pendant les cours.
A la fin de cette description, vous trouverez des illustrations des 10 stades représentés que vous pouvez
également utiliser comme modèle de copie pour vos cours. En décrivant, en inscrivant et en classant les
différents stades correctement, vos élèves peuvent mieux comprendre et approfondir la matière étudiée.
Vous trouverez également des représentations en couleur gratuites sur Internet à l’adresse suivante :
www.3bscientific.com
1. Interphase, stade de la phase G
A l’intérieur de la cellule, le noyau cellulaire ainsi que le nucléole (petit corps sphérique qui se trouve dans
les noyaux cellulaires) (1) et sa membrane nucléaire (2) sont visibles. L’ADN encore non déspiralisé (3) contenant l’information génétique se trouve dans le noyau cellulaire.
La cellule même doit sa stabilité et sa forme à des tubes très fins appelés microtubules (4) qui traversent le
plasma cellulaire. Les microtubules commandent entre autres le mouvement de la cellule et les processus
de transport à l’intérieur de la cellule.
On reconnaît le réticulum endoplasmique (5) dans le plasma cellulaire. Il s’agit d’un système tubulaire
emmêlé servant principalement à la synthèse des lipides, au stockage des ions et à la transformation et
au transport de certaines protéines. Sur la membrane du réticulum endoplasmique rugueux se trouvent
les ribosomes qui synthétisent les protéines qui traversent le réticulum endoplasmique.
On peut également désigner l’appareil de Golgi (6) comme un genre de “glande cellulaire”. Il est constitué
de corps creux, empilés les uns sur les autres (citernes de Golgi) formant de petits vésicules qui se détachent
(vésicules de Golgi) (7). L’appareil de Golgi reçoit des éléments constitutifs de la membrane et des enzymes
du réticulum endoplasmique. Sa tâche principale consiste à collecter, emballer et transporter les sécrétions
et à la formation des lysosomes (vésicules de digestion) (8).
La dégradation des éléments cellulaires (= digestion intracellulaire) constitue la tâche principale
des lysosomes. Les mitochondries (9) sont responsables de la production d'énergie de la cellule.
Les centrioles (10) ont pour tâche de former le fuseau. Il s’agit de cylindres creux formés de tubes
longitudinaux (microtubules).
1
19
Page 20

®
FrançaisMéiose
2. Prophase I
La prophase de la première division de maturation représente la plus longue séquence de la méiose au
point de vue du temps. Au cours de cette phase, les chromosomes et la chromatine modifient leur structure
et leur disposition dans le noyau cellulaire selon un ordre défini. Par conséquent, la prophase I est divisée
en quatre sous-sections (leptotène, zygotène, pachytène et diplotène avec diacinèse). Contrairement à la
prophase mitotique qui dure quelques heures, la prophase I méiotique peut durer des jours, des semaines,
des mois ou des années.
Leptotène
Au commencement de la prophase I (leptotène), le nucléole (corpuscule sphérique présent dans le noyau
cellulaire) (1) et sa membrane nucléaire (2) sont visibles. Les chromosomes (3) sont visibles sous forme de
longs fils fins. Leurs extrémités sont ancrées dans la membrane nucléaire. Chaque chromosome a déjà été
répliqué (dédoublé) au cours de l’interphase et est constitué de deux chromatides-sœurs qui se sont cependant accolées si étroitement que l’on n’arrive pas à les distinguer.
Les centrioles ont également été dédoublés au cours de l’interphase. Les deux paires (4) commencent à
s’éloigner l’une de l’autre en direction des deux pôles cellulaires. Le fuseau central (5), constitué de nombreux microtubules, commence à se former entre ces deux paires.
3. Zygotène et Pachytène
Un homologue maternel (1) et un homologue paternel (2) (composé de chromatides-sœurs) d’une paire de
chromosomes sont représentés en plusieurs couleurs afin de représenter les autres chromosomes (au total
2 x 23).
Zygotène
La phase du zygotène se déclenche aussitôt que les chromosomes homologues commencent à s’accoler
étroitement pour former le complexe synaptonémal (3) (disposition parallèle des partenaires homologues).
La plupart du temps, ce processus commence aux extrémités des chromosomes et se poursuit, de même
qu’une fermeture éclair, jusqu’à l’autre extrémité. L’appariement chromosomique (synapse) s’effectue avec
une haute précision de telle sorte que les gènes correspondants des chromosomes homologues se trouvent
directement les uns en face des autres. Ceci constitue une condition importante de l’échange recombinant
des segments de gènes (enjambement). Chaque paire de chromosomes homologues au cours de la prophase I
méiotique est appelée bivalente. Mais étant donné que chaque chromosome homologue est constitué des
chromatides-sœurs étroitement accolées, on peut également parler de tétrades.
Pachytène
Aussitôt que tous les complexes synaptonémaux sont complètement formés, c’est-à-dire lorsque les chromosomes homologues sont complètement accolés les uns aux autres, la phase du pachytène est atteinte.
A intervalles, des nœuds de recombinaison (4) sont visibles sur les complexes synaptonémaux où a lieu
l’échange des segments de gènes.
4. Diplotène
Après l’échange de quelques segments de gènes, les chromosomes homologues (1) se séparent de plus en
plus les uns des autres, mais restent cependant liés à un ou plusieurs points d’intersection (ponts du chiasma
ou chiasmas) (2). La recombinaison génétique a eu lieu auparavant sur les ponts du chiasma (échange de
l’information héréditaire maternelle et paternelle). Les ovules peuvent rester à l'état du diplotène pendant
des mois, voire des années.
5. Diacinèse
La fin de la prophase I méiotique est déclenchée par le détachement des chromosomes de la membrane
nucléaire (1). Les chromosomes s'épaississent et les chromatides-sœurs, reliées par les centromères
20
Page 21

®
Français Méiose
(séquences courtes, riches en AT de l’ADN) (2), sont visibles. Les chromatides non sœurs dans lesquelles un
échange de segments de gènes a eu lieu sont encore toujours reliées entre elles par des ponts de chiasma (3).
La métaphase I succède à la prophase I. Les phases méiotiques restantes nécessitent moins de 10 % du
temps total d’une méiose complète.
6. Métaphase I
Lors de la transition de la prophase I à la métaphase I, les paires de centrioles (1) sont arrivées aux deux
pôles opposés de la cellule. Un appareil fusorial s'est formé et la membrane nucléaire (2) se dégrade. Les
chromosomes se disposent sur le plan équatorial. Ils forment la plaque de la métaphase. Vus de haut, les
chromosomes apparaissent sous forme d'étoile (monaster ou sphère-mère). Les cinétochores (3) sont des
complexes protéiques déjà formés sur les centromères. Une particularité de la métaphase I méiotique
réside dans le fait que les cinétochores de chaque paire de chromatides-sœurs semblent avoir fusionné.
Les microtubules (4) du fuseau central qui se sont fixés exactement sur les cinétochores de chaque paire
de chromatides-sœurs (5), indiquent par conséquent tous la même direction. Les ponts de chiasma (6)
sont encore toujours préservés. Ils jouent un rôle important pour la disposition correcte des chromosomes
homologues sur le plan équatorial.
Le réticulum endoplasmique (7) et l’appareil de Golgi (8) sont pratiquement décomposés complètement.
7. Anaphase I
Au cours de l’anaphase I de la méiose, les chromosomes homologues (1) se séparent et non les chromatides-sœurs comme au cours de la mitose. Les ponts de chiasma qui retenaient les chromosomes homologues maternels et paternels se décomposent.
Quelques organismes mutants, chez lesquels l’enjambement méiotique n’est effectué que dans une certaine
mesure, présentent des paires de chromosomes exemptes de ponts de chiasma. La plupart du temps, ces
paires ne sont pas vraiment désunies (non-disjonction), et les cellules filles en résultant possèdent un chromosome trop peu ou un chromosome de trop. On désigne une telle déformation par le terme d’aberration
chromosomique numérique, pouvant entraîner des anomalies.
La séparation commence au niveau des cinétochores (2), lieu de fixation des fibres tractrices du fuseau central. C’est à partir de cet endroit que les chromosomes sont lentement tirés par les microtubules (3) se raccourcissant, et par l’effet de traction en résultant vers les centrioles (4) se trouvant aux pôles de la cellule.
Les microtubules (5) qui ne sont pas liés à des chromosomes s’allongent, ce par quoi la distance entre les
centrioles augmente, et la cellule s’étire dans le sens de la longueur. Sur le plan équatorial, on reconnaît
l’ébauche d’un sillon d’étranglement (6).
Par l’enjambement effectué au cours de la prophase et la répartition fortuite des chromosomes maternels
et paternels vers les deux pôles de la cellule, on observe une variation du patrimoine héréditaire (voir
introduction).
8. Télophase I, Cytocinèse I, Intercinèse, Prophase II et Métaphase II
Télophase I et Cytocinèse
Au cours de la télophase I, l’appareil fusorial se résorbe, et un étranglement annulaire (1) se forme au
niveau du plan équatorial. De même, une fine membrane cellulaire (2) se forme. Au cours de la cytocinèse
suivante, le corps cellulaire est divisé en deux parties exactes au centre, c'est-à-dire à l'étranglement annulaire entre les deux nouveaux noyaux fils (3). Chaque noyau fils contient le jeu de chromosomes maternel,
resp. paternel légèrement modifié par l’enjambement, l’ADN est déjà dédoublé, c.-à-d. qu’un chromosome
se compose de deux chromatides-sœurs (4).
21
Page 22

®
FrançaisMéiose
Le réticulum endoplasmique (5) et l’appareil de Golgi (6) ont retrouvé leur forme et leur taille initiales.
A la fin de la cytocinèse, la première division de maturation est achevée.
Intercinèse
La première et la deuxième division de maturation sont séparées par une courte phase de repos (interphase).
Un dédoublement des chromosomes issus de deux chromatides n’a cependant pas lieu (pas de phase S).
Les deux chromatides-sœurs de chaque chromosome restent liées entre elles par les centromères.
Division de maturation II
La deuxième division de maturation se déroule comme une mitose (division habituelle du noyau et de la
cellule). C’est la raison pour laquelle on l’appelle également division équationnelle. Etant donné que les
chromosomes n’ont pas été redédoublés au cours de l'intercinèse précédente, on observe une réduction
du patrimoine héréditaire du jeu de chromosomes haploïde lors de la seconde division méiotique y succédant.
Prophase II
La prophase II correspond étroitement à la prophase d’une mitose et se déroule très rapidement dans
tous les organismes. La perméabilité de la surface de la cellule augmente afin d’absorber le liquide de
l’environnement. L’appareil microtubulaire du cytosquelette se restructure. La membrane nucléaire se
désagrège, et le fuseau se forme à partir de microtubules se restructurant.
Métaphase II
Au cours de la métaphase II, les chromosomes sont à nouveau disposés sur le plan équatorial, et les deux
extrémités du fuseau se trouvent aux deux pôles opposés l’un à l’autre (de façon analogue à la métaphase I).
Une différence essentielle par rapport à la métaphase I réside dans le fait que deux cinétochores se sont
développés sur les chromatides-sœurs qui cette fois indiquent des directions polaires contraires.
9. Anaphase II
Au cours de l’anaphase II, les deux chromatides-sœurs (1) de chaque chromosome sont séparées comme au
cours de la mitose.
La séparation commence au niveau des cinétochores (2), au lieu de fixation des fibres tractrices du fuseau
central. A partir de cet endroit, les chromatides sont tirées lentement par les microtubules (3) se raccourcissant, et par l’effet de traction en résultant vers les centrioles (4) se trouvant aux pôles de la cellule.
Les microtubules (5) qui ne sont pas liés à des chromatides s’allongent, ce par quoi la distance entre les
centrioles augmente, et la cellule s'étire dans le sens de la longueur. Sur le plan équatorial, on reconnaît
l’ébauche d’un sillon d’étranglement (6).
10. Télophase II et Cytocinèse II
De l’étranglement et de la séparation des deux cellules issues de la première division de maturation
résultent maintenant quatre cellules haploïdes (1) possédant une combinaison de gènes différente, causée
par la répartition fortuite des chromosomes et par l’enjambement. Ceci permet d’expliquer le fait que
frères et sœurs ne sont pas identiques : un enfant présente plus les caractéristiques du père, l’autre enfant
présente plus les caractéristiques de la mère. C’est pourquoi des caractéristiques appartenant aux ascendants peuvent réapparaître.
22
Page 23

1
6
7
3
8
2
1
10
4
5
9
2
2
1
3
4
5
4
3
4
3
1
2
23
Page 24

4
5
1
2
1
6
3
8
6
7
2
1
1
5
3
4
2
1
24
Page 25

7
4
6
5
1
2
3
4
8
2
7
1
6
3
4
5
1
2
3
4
6
5
9
25
Page 26

10
1
1
1
1
26
Page 27

®
Português
A meiose, também chamada de divisão reducional, é uma forma especial de divisão celular. Dependendo
do organismo ou do tipo de célula, ela pode durar poucos dias ou também anos. O processo de meiose
origina gametas (células germinativas), ou seja, a divisão reducional de uma célula precursora leva à
formação de quatro gametas. Basicamente, podemos diferenciar dois tipos de gametas. Gametas grandes
e imóveis são chamados de óvulos ou oócitos e gametas pequenos e móveis são chamados de espermatozóides.
Os óvulos são formados nos ovários a partir de células precursoras por meio da divisão meiótica. A maturação dos óvulos do ser humano inicia-se já no estado embrionário (3o a 4o mês de gestação), porém, as
células permanecem em um estado meiótico intermediário até a maturação sexual do organismo. A partir
deste momento, alguns dos óvulos imaturos concluem o processo de divisão meiótica em períodos regulares, controlados por hormônios. Após o início da puberdade, a maturação dos espermatozóides humanos
nos testículos ocorre em intervalos regulares. Neste caso o processo completo da divisão meiótica dura
entre 20 e 24 dias.
Normalmente as células do corpo humano (p. ex. as células precursoras dos gametas) contêm um número
diplóide de cromossomos, ou seja, os cromossomos ocorrem aos pares, sendo um componente do par de
origem paterna e o outro de origem materna. Portanto, cada cromossomo possui uma cópia, sendo parte
de um par de cromossomos correspondentes (homólogos). Os gametas, em contrapartida, são haplóides.
Isto significa que contêm somente um representante de cada par. Portanto, os óvulos ou espermatozóides
possuem apenas a metade dos genes maternos ou paternos; desta maneira, a fusão de ambas as células
leva à formação de uma nova célula (zigoto) com um número de cromossomos diplóide.
O objetivo da meiose consiste em reduzir o número diplóide de cromossomos de uma célula precursora
dos gametas a um número haplóide, a fim de criar as condições prévias para a reprodução sexual. Uma
outra função importante da meiose é a mistura dos genes, obtida através de dois mecanismos:
1. a distribuição aleatória dos cromossomos maternos e paternos às células germinativas formadas;
2. a troca de genes entre os cromossomos homólogos (recombinação genética).
Meiose
(Os respectivos processos destes dois mecanismos serão expostos mais abaixo no âmbito da descrição das
fases individuais.)
No caso do ser humano, que possui um número haplóide de 23 cromossomos, a distribuição aleatória dos
cromossomos resulta em um número de variações genéticas possíveis de 2
de genética é adicionalmente aumentada devido à troca de genes entre os cromossomos.
Antes da divisão meiótica, as células precursoras dos gametas encontram-se no estado da interfase, que
corresponde ao período intermediário entre duas divisões celulares (mitóticas ou meióticas). A interfase
abrange 3 etapas:
• Fase G1 (pré-síntese) Esta fase é caracterizada pelo crescimento da célula.
• Fase S (síntese) Nesta etapa inicia-se a duplicação dos centríolos e do DNA (ácido desoxirribonucléico).
• Fase G2 (pós-síntese) Esta fase marca o fim da síntese de DNA antes da fase de divisão. Além disto, é
concluído o processo de duplicação dos centríolos.
A interfase é seguida pela meiose, que consta de duas fases de divisão separadas por uma curta interfase
especial (intercinese). Como também é o caso na mitose, cada uma destas divisões é subdividida em diferentes etapas:
23
ou 8,4 x 106. Esta variabilida-
27
Page 28

®
PortuguêsMeiose
Divisão I
• Prófase I (abrange quatro subetapas: leptóteno, zigóteno, paquíteno e diplóteno com diacinese)
• Metáfase I
• Anáfase I
• Telófase I
• Citocinese I
Intercinese
Divisão II
• Prófase II
• Metáfase II
• Anáfase II
• Telófase II
• Citocinese II
A série de modelos sobre a meiose da 3B Scientific® (número de produto: R02) e o quadro esquemático da
mitose (V2051M, V2051U) mostra uma célula animal típica em uma escala de aproximadamente 10.000:1.
Na parte inferior dos esquemas encontram-se cortes das organelas celulares.
A série de modelos sobre a meiose da 3B Scientific® é fornecida com uma embalagem equipada com um
dispositivo de suspensão, com o qual é possível pendurar a série de modelos na parede para economizar
espaço. Ímãs fixados no reverso dos modelos possibilitam a sua disposição em quadros magnéticos na sala
de aula.
Ao final desta apresentação, você encontrará esquemas das 10 etapas apresentadas, que podem ser copiados e utilizados em aula. Pintando os esquemas, preenchendo as legendas e colocando as diferentes etapas na ordem certa, seus alunos poderão facilmente entender e aprofundar o que aprenderam.
Esquemas coloridos das diferentes etapas podem ser também gratuitamente obtidos na Internet sob o
endereço www.3bscientific.com.
1. Interfase, fase G
No interior da célula, podemos observar o núcleo celular com o nucléolo (1) e a carioteca (membrana
nuclear) (2). Dentro do núcleo celular encontra-se o DNA ainda não condensado (3) com a informação
genética.
A estabilidade e a forma da célula são garantidos através de cilindros extremamente finos, os chamados
microtúbulos (4) presentes no citoplasma. Os microtúbulos também são responsáveis pelo movimento da
célula e pelos processos de transporte intracelulares.
Dentro do citoplasma podemos observar o retículo endoplasmático (5). Trata-se de uma complexa rede
de tubos, responsável sobretudo pela síntese de lipídios e pelo transporte de íons e certas proteínas no
interior da célula. Junto à membrana do retículo endoplasmático encontram-se os ribossomos, cuja função
consiste na produção de proteínas.
O complexo de Golgi (6) é também chamado de “glândula celular”. Ele é constituído por empilhamentos de
pequenos sacos achatados, em cujas bordas brotam pequenas vesículas (vesículas de Golgi) (7). O complexo
de Golgi recebe enzimas e componentes do retículo endoplasmático para a formação de suas membranas.
Sua função principal consiste no armazenamento e transporte de secreções e na síntese de lisossomos
(vesículas cheias de enzimas digestivas) (8).
A função principal dos lisossomos é a digestão de material celular. Esta digestão pode ocorrer de dentro
para fora da célula (=exocitose) ou dentro da própria célula (=digestão intracelular, endocitose). A energia
dentro da célula é gerada pelas mitocôndrias (9).
O papel dos centríolos (10) consiste em formar o fuso de divisão. Os centríolos têm a forma de cilindros
ocos, formados por túbulos longitudinais (microtúbulos).
28
1
Page 29

®
Português
2. Prófase I
A prófase da primeira divisão é o período de maior duração da meiose. Durante esta etapa, os cromossomos e a cromatina alteram a sua estrutura e sua posição no núcleo conforme uma determinada seqüência.
Por esta razão, a prófase I é subdividida em quatro períodos (leptóteno, zigóteno, paquíteno, diplóteno
com diacinese). Ao contrário da prófase mitótica, que tem uma duração de poucas horas, a prófase I da
meiose pode demorar vários dias, semanas, meses ou anos.
Leptóteno
No início da prófase I (leptóteno), podemos observar o nucléolo (1) e a carioteca (membrana nuclear) (2).
Os cromossomos (3) tornam-se visíveis como filamentos finos, compridos e individualizados, cujas extremidades estão fixadas à carioteca. Os cromossomos replicados (duplicados) durante a interfase precedente
consistem em duas cromátides irmãs que encontram-se muito próximas uma à outra, tornando impossível
a sua distinção.
Os centríolos também são duplicados durante a interfase. Ambos os pares (4) começam a afastar-se gradativamente em direção aos pólos da célula. Entre eles forma-se o chamado fuso meiótico (5), composto por
numerosos microtúbulos.
3. Zigóteno e Paquíteno
O modelo mostra respectivamente um cromossomo homólogo materno (1) e paterno (2) (compostos por
duas cromátides irmãs) de um par de cromossomos, representados em diferentes cores, simbolizando o
conjunto completo de cromossomos (ao todo 2 x 23).
Zigóteno
A etapa do zigóteno inicia-se a partir do momento em que os cromossomos homólogos começarem a se
juntar, formando o complexo sinaptonemático (3) (disposição paralela dos pares homólogos). Em geral,
este processo começa em uma extremidade dos cromossomos e é continuado progressivamente até chegar
à outra extremidade, semelhante a um zíper. Este pareamento dos cromossomos (sinapse) ocorre com
grande precisão, sendo que ao final do processo os genes correspondentes dos cromossomos homólogos
encontram-se diretamente opostos. Este é um pré-requisito para a recombinação de pedaços de genes
(crossing over). Na prófase I da meiose, cada par de cromossomos homólogos é chamado de bivalente;
porém, pelo fato de que cada cromossomo homólogo individual é constituído por duas cromátides irmãs,
o conjunto destes quatro filamentos de cromossomos homólogos pareados é também chamado de tétrade.
Meiose
Paquíteno
Após a formação completa de todos os complexos synaptonemáticos, isto é, após a conclusão do pareamento dos cromossomos homólogos, inicia-se a fase do paquíteno. Nos complexos synaptonemáticos tornam-se visíveis os pontos de recombinação (4), nos quais será realizada a troca de segmentos de genes.
4. Diplóteno
Após as quebras em cromátides homólogas seguidas de soldadura de pedaços de genes trocados, os cromossomos homólogos (1) começam a afastar-se gradativamente, permanecendo em contato em alguns
pontos chamados quiasmas (2). Os quiasmas representam as regiões em que houve a recombinação genética (troca de informação genética materna e paterna). Os óvulos podem permanecer no estado do diplóteno
durante vários meses ou anos.
5. Diacinese
O final da prófase I da meiose inicia-se com a separação dos cromossomos da carioteca (1). A condensação
dos cromossomos aumenta ainda mais e as cromátides irmãs, ligadas através dos centrômeros (seqüências
de DNA curtas e ricas em AT) (2), tornam-se visíveis. As cromátides não irmãs, ou seja, aquelas em que
houve uma troca de segmentos de genes, permanecem em contato nos quiasmas (3).
29
Page 30

®
PortuguêsMeiose
O final da prófase I marca o início da metáfase I. As fases meióticas restantes têm uma duração de menos
de 10% da duração total de uma meiose completa.
6. Metáfase I
No final da prófase I e no início da metáfase I, os pares de centríolos (1) chegaram aos dois pólos opostos
da célula. Formou-se um fuso e a carioteca (2) se dissolve. Os cromossomos se organizam no plano equatorial
da célula. Eles formam a chamada placa metafásica (placa equatorial). Vistos de cima, os cromossomos
apresentam um aspecto de estrela (monáster ou estrela-mãe). Os cinetócoros (3) são complexos de proteína
que se formaram nos centrômeros. Um aspecto específico da metáfase I da meiose é o fato de os cinetócoros
de cada par de cromátides irmãs parecerem estar fusionados. Desta maneira, os microtúbulos (4) do fuso
meiótico, que se ligaram exatamente aos cinetócoros de cada par de cromátides irmãs (5), apontam todos
para a mesma direção. Os quiasmas (6) ainda permanecem existentes. Eles desempenham um papel central
para a organização correta dos cromossomos homólogos no plano equatorial.
O retículo endoplasmático (7) e o complexo de Golgi (8) estão quase completamente desintegrados.
7. Anáfase I
Durante a anáfase I da meiose ocorre a separação dos cromossomos homólogos (1) e não, como é o caso
na mitose, das cromátides irmãs. Os quiasmas, que até este momento haviam ligado os cromossomos
homólogos maternos e paternos, desaparecem.
Alguns organismos mutantes, em que o crossing over meiótico ocorre somente em medida limitada,
apresentam pares de cromossomos sem quiasmas. Em geral, estes pares não são separados corretamente
(nondisjunction) e as células-filhas resultantes contêm um cromossomo a mais ou a menos. Uma mutação
deste tipo é chamada de aberração cromossômica numérica, que resulta em malformações.
A separação inicia-se nos cinetócoros (2), os pontos aos quais se prendem as fibras do fuso de divisão.
Através do encurtamento dos microtúbulos (3) e o efeito de tração resultante, os cromossomos migram
em direção aos centríolos (4) localizados nos pólos da célula. Os microtúbulos (5) que não estão ligados a
cromossomos tornam-se mais longos, aumentando a distância entre os centríolos e tornando a célula mais
longa. No plano equatorial da célula começa a aparecer uma constrição (6).
Devido ao crossing over na pró-fase e a distribuição aleatória dos cromossomos maternos e paternos aos
pólos da célula é atingida uma grande variabilidade genética (veja introdução).
8. Telófase I, Citocinese I, Intercinese, Prófase II e Metáfase II
Telófase I e Citocinese
Durante a telófase I, o fuso começa a desaparecer e uma constrição (1) forma-se no plano equatorial. Além
disto, é formada uma carioteca fina (2). Durante a citocinese que segue, o corpo celular é dividido exatamente
ao meio, na constrição entre os dois núcleos filhos recém-formados (3). Através do crossing over, os
núcleos-filhos recebem cromossomos maternos e paternos diferentes, sendo que o DNA já foi duplicado,
isto é, um cromossomo é composto por duas cromátides irmãs (4).
O retículo endoplasmático (5) e o complexo de Golgi (6) voltam a adquirir a sua forma e seu tamanho original.
O final da citocinese corresponde ao fim da primeira divisão.
Intercinese
A primeira e a segunda divisão são separadas por uma curta fase de repouso (interfase). Porém, não ocorre
uma duplicação dos cromossomos compostos por duas cromátides (não há uma fase S). As cromátides
30
Page 31

®
Português Meiose
irmãs de cada cromossomo permanecem ligadas através dos centrômeros (7).
Divisão II
A segunda etapa de divisão corresponde ao processo da mitose (divisão celular e nuclear normal). Por esta
razão, ela é também chamada de divisão equacional. Sendo que os cromossomos não foram novamente
duplicados durante a intercinese, a segunda divisão meiótica resulta na redução dos genes ao número
haplóide de cromossomos.
Prófase II
A prófase II é uma prófase semelhante à da mitose e é muito rápida em todos os organismos. A permeabilidade
da membrana celular aumenta, possibilitando a absorção de líquidos. O complexo de microtúbulos do
citoesqueleto é reorganizado. A carioteca desaparece e o fuso é novamente formado a partir dos microtúbulos reorganizados.
Metáfase II
Durante a metáfase II, os cromossomos se posicionam novamente no plano equatorial da célula e as duas
extremidades do fuso migram para os pólos opostos da célula (igual à metáfase I). Uma diferença importante em relação à metáfase I consiste no fato de que nas cromátides irmãs se desenvolveram dois cinetócoros, que agora estão orientados para pólos diferentes, em direção oposta.
9. Anáfase II
Segue a anáfase II, durante a qual as duas cromátides irmãs (1) de cada cromossomo são separadas, como
é também o caso na mitose.
A separação inicia-se nos cinetócoros (2), aos quais se prendem as fibras do fuso de divisão. Através do
encurtamento dos microtúbulos (3) e o efeito de tração resultante, os cromossomos são arrastados gradualmente em direção aos centríolos (4) localizados nos pólos da célula. Os microtúbulos (5) que não estão ligados a cromátides tornam-se mais compridos, aumentando assim a distância entre os centríolos e tornando
a célula mais longa. No plano equatorial, começa a aparecer uma constrição (6).
10. Telófase II e citocinese II
Através da constrição e divisão das duas células resultantes da primeira divisão meiótica formam-se quatro
células haplóides (1) com diferentes combinações genéticas causadas pela distribuição aleatória dos
cromossomos e pelo crossing over. Isto explica o fato de irmãos nunca serem idênticos: um filho apresenta
mais características do pai, o outro tem a aparência da mãe. E desta maneira também podem reaparecer
características de outros parentes antepassados.
31
Page 32

®
Meiosi
La meiosi, o divisione riduzionale, è un tipo particolare di divisione cellulare. A seconda dell’organismo e
della varietà di cellula interessata, può durare da alcuni giorni ad alcuni anni e ha come risultato la produzione di cellule sessuali (gameti). Ogni cellula precursore produce quattro gameti attraverso la divisione
riduzionale. In generale, esistono due tipi di gameti. Le grandi cellule immobili vengono definite cellule
uovo od ovociti, mentre i gameti piccoli e mobili sono gli spermatozoi o spermatociti.
Le cellule uovo vengono create attraverso la divisione meiotica da parte di cellule precursori nelle ovaie.
Benché le cellule uovo umane inizino a maturare già nell’embrione (dal 3° all’8° mese di gravidanza),
esse restano in una fase meiotica intermedia fino al raggiungimento della maturità sessuale. Da questo
momento in poi, alcune cellule uovo immature completano la divisione meiotica a intervalli regolari sotto
il controllo degli ormoni. La maturazione degli spermatozoi umani avviene regolarmente nei testicoli una
volta raggiunta la maturità sessuale. In questo caso, una divisione meiotica completa richiede 20-24 giorni.
Di solito, le cellule somatiche, cioè i precursori dei gameti, contengono due assetti cromosomici e vengono
perciò definite diploidi: metà dei cromosomi deriva dalla madre e l‘altra metà dal padre. Di conseguenza,
esiste una copia esatta di ciascun cromosoma, ossia, una coppia corrispondente (omologa) di cromosomi.
Al contrario, i gameti contengono un singolo assetto cromosomico e vengono quindi definiti aploidi. In
altre parole, le cellule uovo e gli spermatozoi contengono solo metà del patrimonio genetico della madre e
del padre, così che quando si uniscono, danno origine a una nuova cellula (zigote) che presenta un assetto
cromosomico diploide completo.
Lo scopo della meiosi è ridurre l‘assetto cromosomico normalmente diploide di una cellula precursore
di un gamete trasformandolo in un assetto aploide, per stabilire così la base della riproduzione sessuale.
Un‘altra importante funzione della meiosi è di mischiare le informazioni genetiche. Tale obiettivo viene
raggiunto tramite due meccanismi:
1. una distribuzione casuale dei cromosomi materni e paterni alle cellule sessuali che vengono prodotte
2. lo scambio dei geni tra i cromosomi omologhi (ricombinazione genetica)
EnglishItaliano
Le procedure alla base di questi due meccanismi sono spiegate più avanti nella descrizione delle singole
fasi.
Negli umani, che dispongono di 23 cromosomi nell’assetto aploide, la sola distribuzione casuale dei cromosomi rende possibili 223, ossia 8,4 x 106 diverse varianti genetiche. Il numero di varianti aumenta ulteriormente grazie allo scambio di geni tra i cromosomi.
Prima della divisione meiotica, le cellule precursori dei gameti si trovano nell‘interfase, che si riferisce al
periodo tra due divisioni cellulari (mitotica o meiotica). L’interfase comprende tre stadi:
• Fase G
• Fase S (sintesi), in cui i centrioli e il DNA (acido desossiribonucleico) iniziano a duplicarsi.
• Fase G2 (lacuna postsintetica), che separa la conclusione della sintesi del DNA dalla fase di divisione.
Viene inoltre completata la duplicazione dei centrioli.
La meiosi, stadio che segue l’interfase, comprende due successive divisioni (meiotiche) di maturazione,
separate da una breve interfase specifica (intercinesi). Come nella mitosi, anche nella divisione meiotica si
riconoscono diversi stadi:
(lacuna presintetica), ossia la fase in cui la cellula cresce.
1
32
Page 33

®
EnglishItaliano Meiosi
Prima divisione meiotica:
• Profase I (quattro sottostadi: leptotene, zigotene, pachitene e diplotene con diacinesi)
• Metafase I
• Anafase I
• Telofase I
• Citocinesi I
Intercinesi
Seconda divisione meiotica:
• Profase II
• Metafase II
• Anafase II
• Telofase II
• Citocinesi II
La serie di modelli 3B Scientific® sulla meiosi (codice prodotto R02) e il tabellone murale sulla meiosi
(V2051M, V2051U) mostrano una tipica cellula mammifera con un fattore di ingrandimento pari a circa
10.000. Nella parte inferiore dei modelli e delle illustrazioni, gli organelli cellulari vengono visualizzati
come se fossero aperti.
La serie viene fornita in un supporto per il deposito, dotato di un dispositivo che permette di appendere i
modelli al muro, così da risparmiare spazio. Questi ultimi sono inoltre dotati di calamite sul retro che permettono di applicarli e disporli su lavagne magnetiche in classe e utilizzarli a fini educativi.
Al termine di questa descrizione seguono le illustrazioni delle 10 fasi, che possono essere fotocopiate per
le lezioni. Colorando, denominando e ordinando le singole fasi nel modo corretto, gli studenti potranno
facilmente ripassare e memorizzare quanto appreso.
Illustrazioni a colori gratuite dei vari stadi sono disponibili anche su Internet all‘indirizzo www.3bscientific.
com.
1. Interfase, stadio della fase G
All’interno della cellula è possibile scorgere il nucleo con il nucleolo (1) e la membrana nucleare (2). Il
nucleo contiene inoltre il DNA (3) non ancora elicoidale con le informazioni genetiche.
La cellula riceve la propria stabilità e forma da tubi molto sottili, i cosiddetti microtubuli (4), che si estendono attraverso il citoplasma. Essi controllano, tra le altre cose, i movimenti della cellula e i processi di
trasporto intracellulari.
All’interno del citoplasma è visibile il reticolo endoplasmatico (5). Esso è costituito da un sistema di tubi
intrecciati e destinati alla sintesi dei lipidi, all‘immagazzinamento di ioni, alla trasformazione e al trasporto di determinate proteine. La membrana del reticolo endoplasmatico rugoso è ricca di ribosomi, la cui
funzione è sintetizzare le proteine che attraversano il reticolo stesso.
L’apparato del Golgi (6) può essere definito la “ghiandola della cellula”. È costituito da una serie di sacche
cave impilate (cisterne membranose appiattite) che si gonfiano man mano che le vescicole del Golgi (7) si
rimpiccioliscono e si sciolgono. L’apparato del Golgi riceve componenti membranosi ed enzimi dal reticolo
endoplasmatico: la sua funzione principale è di raccogliere e distribuire secrezioni, oltre che di produrre
lisosomi (vescicole digestive) (8).
Lo scopo principale dei lisosomi è disgregare i componenti della cellula (digestione intracellulare).
1
33
Page 34

®
ItalianoMeiosi
Gli organelli addetti alla produzione di energia sono i mitocondri (9).
La funzione dei centrioli (10) è assemblare il fuso mitotico. Si tratta di cilindri cavi costituiti da tubi disposti
longitudinalmente (microtubuli).
2. Profase I
La profase della prima divisione meiotica è la parte del processo che richiede più tempo. Nel corso del
suo svolgimento, i cromosomi e la cromatina cambiano la propria struttura e disposizione all‘interno del
nucleo, seguendo un ordine specifico. Di conseguenza, la profase I è suddivisa in quattro sottostadi (leptotene, zigotene, pachitene e diplotene con diacinesi). Al contrario della profase mitotica, che dura diverse
ore, la profase meiotica I può durare giorni, settimane, mesi o anni.
Leptotene
All’inizio della profase I (leptotene), è possibile riconoscere il nucleolo (1) e la membrana nucleare (2).
I cromosomi (3) sono visibili sotto forma di individuali filamenti lunghi e sottili. Le loro estremità sono
collegate all’interno della membrana nucleare. Ogni cromosoma è già stato replicato, ossia duplicato, nel
corso dell‘interfase ed è costituito da due cromatidi fratelli, che sono tuttavia talmente vicino l‘uno all‘altro
che non è possibile differenziarli. Anche i centrioli sono stati duplicati durante l’interfase. Entrambe le
coppie (4) iniziano a muoversi in direzioni opposte verso i due poli della cellula. Tra di essi si forma ciò che
viene definito fuso centrale (5), costituito da numerosi microtubuli.
3. Zigotene e pachitene
Un omologo materno (1) e uno paterno (2), costituiti da due cromatidi fratelli, di una coppia di cromosomi
sono visualizzati con colori diversi per rappresentare gli altri cromosomi (2 x 23 in totale).
Zigotene
Lo zigotene inizia non appena i cromosomi omologhi cominciano ad allinearsi gli uni accanto agli altri per
formare il complesso sinaptinemale (3), con una disposizione parallela a quella dei loro partner omologhi.
Questo processo si verifica solitamente a partire da un‘estremità dei cromosomi e continua fino all’altra
estremità, in modo analogo a una chiusura lampo. L’appaiamento dei cromosomi (sinapsi) avviene in
modo molto preciso, in modo che i geni corrispondenti dei cromosomi omologhi siano rivolti uno direttamente di fronte all‘altro. Questo è un requisito importante per lo scambio ricombinante delle sezioni genetiche (crossing-over). Le coppie di cromosomi omologhi nella profase meiotica I vengono di norma definite
bivalenti, ma poiché ciascun cromosoma omologo è costituito da cromatidi fratelli vicini gli uni agli altri,
possono essere chiamate anche tetradi.
Pachitene
Il pachitene inizia non appena tutti i complessi sinaptinemali sono completamente sviluppati, ossia i cromosomi omologhi sono allineati. Da questo momento in poi, sui complessi sinaptinemali, dove avviene lo
scambio delle sezioni genetiche, i noduli di ricombinazione (4) diventano visibili a intervalli.
4. Diplotene
Dopo lo scambio di alcune sezioni genetiche, i cromosomi omologhi (1) si separano sempre più, restando
collegati in corrispondenza di uno o più punti del crossing-over (chiasmi) (2). I chiasmi cromosomici sono
i punti in cui è avvenuta in precedenza la ricombinazione genetica, ossia lo scambio delle informazioni
genetiche materne e paterne. Le cellule uovo possono restare nello stadio diplotene per mesi, o addirittura
per anni.
5. Diacinesi
La profase meiotica I volge al termine quando i cromosomi si staccano dalla membrana nucleare (1). I
cromosomi sono condensati e diventano visibili i cromatidi fratelli, uniti dal centromero (brevi sequenze di
DNA con un elevato livello di AT)(2). I cromatidi non fratelli in cui è avvenuto uno scambio di sezioni genetiche restano collegati per mezzo dei chiasmi (3).
34
Page 35

®
Italiano Meiosi
La fase successiva alla profase I è la metafase I. Le fasi meiotiche rimanenti a questo punto richiedono
meno del 10% del tempo totale necessario per una meiosi completa.
6. Metafase I
Durante la transizione dalla profase I alla metafase I, le coppie di centrioli (1) hanno raggiunto i due poli
opposti della cellula. Si è sviluppato un apparato fusolare e la membrana nucleare (2) si dissolve. I cromosomi si allineano a livello equatoriale, formando la cosiddetta piastra metafasica. Visti dall’alto assumono
una forma simile a una stella (monaster o stella “madre”). I cinetocori (3) sono complessi di proteine già
sviluppati in corrispondenza dei centromeri. Una particolarità della metafase meiotica I è che i cinetocori
di ciascuna coppia di cromatidi fratelli sembrano essersi uniti. I microtubuli (4) del fuso centrale, che sono
ora collegati in modo preciso ai cinetocori di ciascuna coppia di cromatidi fratelli (5), puntano di conseguenza tutti nella stessa direzione. I chiasmi (6) sono ancora intatti. Esso ricoprono un ruolo importante
nel corretto allineamento dei cromosomi omologhi a livello equatoriale.
Il reticolo endoplasmatico (7) e l’apparato del Golgi (8) sono quasi completamente dissolti.
7. Anafase I
Nell’anafase I della meiosi, sono i cromosomi omologhi (1) a separarsi, invece dei cromatidi fratelli, come
avviene invece nella mitosi. Durante il processo si dissolvono i chiasmi, che fino a questo momento avevano tenuto uniti i cromosomi materni e paterni.
Alcuni organismi mutanti, dove il crossing-over meiotico avviene solo entro certi limiti, presentano coppie
di cromosomi senza chiasmi. Di solito, queste coppie non sono completamente separate (non-disgiunzione)
e le cellule figlie risultanti hanno un cromosoma in meno o in più. Tali malformazioni vengono definite
aberrazioni cromosomiche numeriche e causano deformazioni.
La disgiunzione inizia in corrispondenza dei cinetocori (2), punto dove sono collegate le fibre del fuso centrale. Da qui, i cromosomi vengono trascinati lentamente verso i centrioli (4), collocati ai poli della cellula,
spostandosi lungo i microtubuli (3) che creano un effetto di trazione man mano che si accorciano. I microtubuli (5) non collegati ai cromosomi si allungano, aumentando così la distanza tra i centrioli ed estendendo la cellula. A livello equatoriale, diventa visibile lo stadio iniziale di un solco di clivaggio (6).
Il processo di crossing-over durante la profase e la distribuzione casuale dei cromosomi materni e paterni
ai poli della cellula portano a una diversificazione delle informazioni genetiche (vedere l‘introduzione).
8. Telofase I, citocinesi I, intercinesi, profase II e metafase II
Telofase I e citocinesi
Nella telofase I, il fuso si disintegra e a livello equatoriale si sviluppa una strozzatura (1). Viene inoltre a
crearsi una sottile membrana nucleare (2). Nel corso della successive fase di citocinesi, il corpo della cellula
si divide esattamente al centro, in corrispondenza della strozzatura tra i nuclei delle due nuove cellule
figlie (3). I nuclei figli contengono ciascuno l’assetto cromosomico materno e paterno leggermente alterato
attraverso il processo di crossing-over, dove il DNA è già presente in forma duplicata, ossia un cromosoma è
costituito da due cromatidi fratelli (4).
Il reticolo endoplasmatico (5) e l’apparato del Golgi (6) hanno riassunto la loro forma e le loro dimensioni
iniziali.
Alla fine della citocinesi, la prima divisione meiotica è completa.
35
Page 36

®
ItalianoMeiosi
Intercinesi
La prima e la seconda divisione meiotica sono separate da un breve periodo di riposo (interfase). Tuttavia,
non avviene alcuna duplicazione dei cromosomi costituiti da due cromatidi (non è presente alcuna fase S).
Entrambi i cromatidi fratelli di ciascun cromosoma restano collegati dai centromeri (7).
Divisione meiotica II
La seconda divisione meiotica avviene esattamente come la mitosi (normale divisione nucleare e cellulare).
Viene perciò anche definita divisione equazionale. Poiché i cromosomi non sono stati nuovamente duplicati durante la precedente intercinesi, la seconda divisione meiotica che ora segue include la riduzione
delle informazioni genetiche al solo assetto cromosomico aploide.
Profase II
La profase II è molto simile a quella della mitosi e avviene molto rapidamente in tutti gli organismi. La
permeabilità della superficie cellulare aumenta al fine di consentire l’assorbimento di liquido dall’esterno.
Il complesso microtubulare del citoscheletro si riorganizza, mentre la membrana nucleare si dissolve e il
fuso viene creato riordinando i microtubuli.
Metafase II
Nella metaphase II, I cromosomi vengono nuovamente posizionati a livello equatoriale e le due estremità
del fuso collocate ai due poli opposti della cellula, come nella metafase I. Una differenza sostanziale
rispetto a quest’ultima è che i due cinetocori si sono sviluppati in corrispondenza dei cromatidi fratelli, i
quali, in questo caso, puntano verso direzioni polari opposte.
9. Anafase II
Durante l’anafase II, i due cromatidi fratelli (1) di ciascun cromosoma vengono disgiunti, proprio come
nella mitosi.
La separazione inizia in corrispondenza dei cinetocori (2), punto dove sono collegate le fibre del fuso centrale. Da qui, i cromosomi vengono trascinati lentamente verso i centrioli (4), collocati ai poli della cellula,
spostandosi lungo i microtubuli (3) che creano un effetto di trazione man mano che si accorciano. I microtubuli (5) non collegati ai cromatidi si allungano, aumentando così la distanza tra i centrioli ed estendendo
la cellula. A livello equatoriale, diventa visibile lo stadio iniziale di un solco di clivaggio (6).
10. Telofase II e citocinesi II
Il clivaggio e la divisione delle due cellule prodotte durante la prima fase meiotica portano ora alla produzione di quattro cellule aploidi (1) con diverse combinazioni genetiche derivanti da una distribuzione cromosomica casuale e dal crossing-over. Ciò spiega perché i fratelli non sono identici: un bambino può avere
più tratti del padre, l’altro della madre. È inoltre possibile che riappaiano caratteristiche degli antenati.
36
Page 37

®37®38®39®40®41®
Page 38

Page 39

Page 40

Page 41

Page 42

Мейоз
Мейоз или редукционное деление - это особый тип клеточного деления. В зависимости от организма и
типа клеток он может длиться от нескольких дней до нескольких лет, приводя к образованию половых
клеток (гамет). Каждая клетка-предшественница гаметы образует четыре гаметы за один цикл
редукционного деления. Всего существует два типа гамет. Крупные, неподвижные клетки называются
яйцеклетки или ооциты, а мелкие, подвижные гаметы - сперматозоиды или сперматоциты.
Яйцеклетки образуются в яичниках в результате мейотического деления из клеток-предшественников.
Яйцеклетки человека начинают созревать уже у эмбриона (3 – 8 месяц беременности), однако эти клетки
остаются в промежуточной фазе мейоза до момента достижения полового созревания. Тогда под
контролем гормонов некоторые незрелые яйцеклетки подвергаются мейотическому делению с
регулярными интервалами. Созревание сперматозоидов человека регулярно происходит в яичках после
достижения половой зрелости. В этом случае полный цикл мейотического деления составляет 20-24 дня.
Обычно соматические клетки тела (например, клетки-предшественники гамет) содержат удвоенный
(диплоидный) набор хромосом, одна половина которого происходит от матери, а другая половина от
отца. Поэтому существует удвоенная копия каждой хромосомы, т.е. совместимая (гомологичная) пара
хромосом. В противоположность этому, гаметы содержат только один (гаплоидный) набор хромосом.
Другими словами, яйцеклетки или сперматозоиды содержат только половину генетической информации
матери и отца, таким образом при слиянии обеих клеток может образоваться новая клетки (зигота) с
полным диплоидным набором хромосом.
Следовательно, целью мейоза является редукция нормального диплоидного набора хромосом клеткипредшественницы гаметы до гаплоидного набора с целью создания основы для любого полового
размножения. Дополнительной важной функцией мейоза является смешение генетической информации.
Это достигается с помощью двух механизмов:
1. случайное распределение материнских и отцовских хромосом в образующихся половых клетках;
2. обмен генами между гомологичными хромосомами (генетическая рекомбинация).
(Процессы, лежащие в основе обоих механизмов, объясняются ниже в описании отдельных фаз).
У людей, которые имеют 23 хромосомы в гаплоидном наборе, случайное распределение хромосом
позволяет создать 1 на 223, т.е. 8,4 х 106 различных возможных генетических вариаций. Кроме того,
количество вариаций возрастает при обмене генами между хромосомами.
До начала мейотического деления клетки-предшественницы гамет находятся в интерфазе, которая
соответствует периоду между двумя (митотическим или мейотическим) циклами клеточного деления.
Интерфаза включает три стадии:
Русский
• Фаза G1 (пресинтетическая) Стадия роста клетки.
• Фаза S (синтетическая) Во время этой фазы начинается удвоение центриолей и ДНК
(дезоксирибонуклеиновой кислоты).
• Фаза G2 (постсинтетическая) Эта фаза отделяет окончание синтеза ДНК от фазы деления. Кроме
того, происходит удвоение центриолей.
Мейоз - фаза, следующая за интерфазой, включает два последовательных деления созревания
(мейотических), которые разделяются короткой специфической отдельной интерфазой (интеркинез).
Как и в ходе митоза, в каждом мейотическом делении выделяют несколько стадий деления:
42
Page 43

®
DeutschРусский
Первое мейотическое деление:
• Профаза I (четыре подраздела: лептотена, зиготена, пахитена и диплотена с диакинезом)
• Метафаза I
• Анафаза I
• Телофаза I
• Цитокинез I
Интеркинез
Второе мейотическое деление:
• Профаза II
• Метафаза II
• Анафаза II
• Телофаза II
• Цитокинез II
Серия моделей 3B Scientific® для изучения мейоза (номер продукта R02) и настенная схема мейоза
(V2051M, V2051U) демонстрируют типичную клетку млекопитающего в увеличении около 10000 раз.
В нижней трети моделей/иллюстраций видны клеточные органеллы в «открытой» клетке.
Серия моделей 3B Scientific® для изучения мейоза поставляется с системой хранения, которая оборудована
подвесным устройством. Серию моделей можно просто повесить на стену в целях экономии пространства.
На обратной стороне моделей также имеются магниты, чтобы в целях обучения их можно было расположить
на магнитных досках в классах.
В конце данного описания Вы найдете иллюстрации 10 фаз мейоза. Вы можете использовать их чтобы
изготовить фотокопии для Ваших занятий. Раскрашивая, подписывая и правильно располагая отдельные
фазы, Ваши студенты смогут легко наглядно изучить и запомнить пройденный материал.
Цветные иллюстрации отдельных стадий также есть в свободном доступе в Интернете на сайте:
www.3bscientific.com.
Мейоз
1. Интерфаза (стадия фазы G1)
Внутри клетки можно увидеть ядро с ядрышком (1) и его ядерную мембрану (2). Ядро также содержит
еще не закрученную в спираль нить ДНК (3) с генетической информацией.
Собственно клетка поддерживает стабильность и форму благодаря очень тонким трубкам, так называемым
микротрубочкам (4), протянутым через цитоплазму. Кроме всего прочего, микротрубочки контролируют
движения клетки и процессы внутриклеточного транспорта.
В цитоплазме можно увидеть эндоплазматический ретикулум (5). Это система переплетенных трубочек,
которая главным образом отвечает за синтез липидов, хранение ионов и реконструкцию и транспортировку
определенных белков. К мембране грубого эндоплазматического ретикулума прикреплены рибосомы,
которые синтезируют белки, передвигающиеся по эндоплазматическому ретикулуму.
Комплекс (или аппарат) Гольджи (6) называемый также «клеточной железой». Он состоит из «стопок»
слоистых полых мешочков (цистерны Гольджи), которые выбухают в виде пузырьков и «отшнуровываются»
(пузырьки Гольджи) (7). В аппарат Гольджи поступают компоненты мембраны и ферменты из
эндоплазматического ретикулума. Главная его функция – накопление, упаковка и транспортировка
секретов и продукция лизосом (переваривающих пуз) (8).
Главной функцией лизосом является разрушение клеточных компонентов (= внутриклеточное переваривание).
Митохондрии (9) отвечают за продукцию энергии для клетки.
Функцией центриолей (10) является образование веретена деления. Они представляют собой полые
цилиндры, состоящие из продольно расположенных трубочек (микротрубочек).
43
Page 44

®
РусскийМейоз
2. Профаза I
Профаза первого мейотического деления является самой продолжительной частью мейоза. Во время этой
фазы хромосомы и хроматин изменяют свою структуру и располагаются в ядре в определенном порядке.
Поэтому профаза I подразделяется на четыре подраздела (лептотена, зиготена, пахитена и диплотена с
диакинезом). В противоположность митотической профазе, которая длится несколько часов, мейотическая
профаза I может продолжаться несколько дней, недель, месяцев или лет.
Лептотена
В начале профазы I (лептотена) можно увидеть ядро (1) и ядерную мембрану (2). Хромосомы (3) теперь
видны как отдельные, длинные, тонкие нити. Их концы прикрепляются к ядерной мембране. Каждая
хромосома уже прошла процесс репликации (удвоения) во время интерфазы и состоит из двух
сестринских хроматид, которые однако так тесно прилежат друг к другу, что их нельзя различить.
Центриоли также удваиваются в интерфазе. Обе пары (4) начинают двигаться к двум полюсам клетки в
противоположных направлениях. Между ними начинает образовываться так называемое центральное
веретено (5), которое состоит из множества микротрубочек.
3. Зиготена и пахитена
Одна материнская (1) и одна отцовская гомологичная хромосома (2) (состоящие из двух сестринских
хроматид) одной пары хромосом показаны разными цветами, чтобы представить другие хромосомы
(всего 2 х 23).
Зиготена
Как только гомологичные хромосомы начинают выстраиваться в линию бок о бок для формирования
синаптонемального комплекса (3) (параллельное расположение гомологичных партнеров), начинается
фаза зиготены. Этот процесс обычно начинается на концах хромосом и продолжается вниз к другому
концу по принципу застежки-молнии. Образование пар хромосом (синапсис) происходит с высокой
точностью, так что совместимые гены гомологичных хромосом сразу располагаются друг против друга.
Это является важным условием для рекомбинантного обмена участками генов (кроссинговер).
Гомологичные пары хромосом в мейотической профазе I обычно называются бивалентами, но поскольку
каждая гомологичная хромосома состоит из тесно соприкасающихся сестринских хроматид, их можно
также назвать тетрадами.
Пахитена
После того, как все синаптонемальные комплексы полностью образованы, т.е. все гомологичные
хромосомы выстроены в линию, начинается фаза пахитены. Теперь становятся видимыми точки
рекомбинации (4), расположенные через интервалы на синаптонемальных комплексах, где
происходит обмен участками генов.
4. Диплотена
После обмена несколькими участками генов гомологичные хромосомы (1) все больше и больше
расцепляются, оставаясь связанными в одной или более точках кроссинговера (мостики хиазмы или
хиазмата) (2). Мостики хиазмы – это участки, где генетическая рекомбинация (обмен материнской и
отцовской генетической информацией) происходит раньше. Яйцеклетки могут оставаться в фазе
диплотены в течение нескольких месяцев или даже лет.
5. Диакинез
Завершение мейотической профазы I начинается, когда хромосомы отсоединяются от ядерной
мембраны (1). Хромосомы конденсируются и становятся видимыми сестринские хроматиды,
связанные центромерами (короткие последовательности ДНК с высоким уровнем АТ) (2).
Не сестринские хроматиды, в которых произошел обмен участками генов, остаются связанными посредством
44
Page 45

®
DeutschРусский
мостиков хиазмы (3).
За профазой I следует метафаза I. Фаза мейоза, описанная до сих пор, занимает менее 10% всего
времени, которое требуется для полного цикла мейоза.
6. Метафаза I
Во время перехода из профазы I в метафазу I пары центриолей (1) достигают двух противоположных
полюсов клетки. Образовалось веретено деления и растворилась ядерная мембрана (2). Хромосомы
выстраиваются в области экватора, образуя так называемую метафазную пластинку. При виде со
стороны полюсов хромосомы имеют форму звезды (монастер или «материнская» звезда). Кинетохоры (3)
- это белковые комплексы, которые уже образовались в центромерах. Особенностью мейотической
метафазы I является то, что кинетохоры каждой пары сестринских хроматид кажутся слившимися.
Микротрубочки (4) центрального веретена, которые теперь тщательно прикреплены к кинетохорам
каждой пары сестринских хроматид (5), все повернуты в одном направлении. Мостики хиазмы (6) еще
не повреждены. Они играют важную роль в правильном выстраивании гомологичных хромосом в области
экватора.
Эндоплазматический ретикулум (7) и аппарат Гольджи (8) теперь почти полностью растворились.
7. Анафаза I
В анафазе I мейоза гомологичные хромосомы (1) разъединяются на сестринские хроматиды, но не так,
как в митозе. В ходе этого процесса растворяются мостики хиазмы, которые так долго удерживали вместе
материнские и отцовские хромосомы.
У некоторых видоизмененных организмов, у которых процесс кроссинговера происходит только в
ограниченной степени, пары хромосом не имеют мостиков хиазмы. Эти пары обычно не полностью
расходятся (неразобщенные), и в результате дочерние клетки имеют на одну хромосому меньше или
больше. Такие дефекты относятся к количественным хромосомным аберрациям, которые вызывают
пороки развития.
Расхождение начинается в кинетохорах (2) – месте, где прикрепляются тянущие нити центрального веретена.
Отсюда хромосомы медленно растягиваются по направлению к центриолям (4), расположенным на
полюсах клетки, двигаясь вдоль микротрубочек (3), которые создают эффект тяги за счет сокращения.
Микротрубочки (5), которые не соединены с хромосомами, теперь становятся длиннее, таким образом
увеличивая расстояние между центриолями и удлиняя клетку. В области экватора становится видна
начинающаяся стадия образования борозды деления (6).
Процесс кроссинговера в ходе профазы и случайное распределение материнских и отцовских хромосом
к полюсам клетки приводит к разнообразию генетической информации (см. введение).
Мейоз
8. Телофаза I, цитокинез I, интеркинез, профаза II и метафаза II
Телофаза I и цитокинез
В телофазе I разрушается веретено и на уровне экватора образуется кольцо сокращения (1). Кроме того,
образуется тонкая ядерная мембрана (2). Во время следующей фазы цитокинеза тело клетки разделяется
точно посередине, в кольце сокращения между двумя новыми дочерними ядрами (3). Каждое дочернее
ядро содержит материнский или отцовский набор хромосом, немного измененный в процессе кроссинговера,
с уже удвоенным количеством ДНК, т.е. одна хромосома состоит из двух сестринских хроматид (4).
Эндоплазматический ретикулум (5) и аппарат Гольджи (6) возвратились к своим начальным форме и
размеру.
В конце цитокинеза заканчивается первое мейотическое деление.
45
Page 46

®
РусскийМейоз
Интеркинез
Первое и второе деление мейоза разделяются коротким периодом покоя (интерфаза). Однако удвоения
хромосом, состоящих из двух хроматид, не происходит (нет S фазы). Обе сестринские хроматиды каждой
хромосомы остаются связанными центромерами (7).
Деление мейоза II
Второе деление мейоза происходит так же, как митоз (обычное ядерное и клеточное деление).
Поэтому оно также называется эквационным делением. Поскольку хромосомы не были вновь удвоены в
ходе предшествующего интеркинеза, второе мейотическое деление, происходящее в это время, включает
уменьшение генетической информации до гаплоидного набора хромосом.
Профаза II
Профаза II в основном подобна профазе митоза и происходит очень быстро у всех организмов.
Проницаемость поверхности клетки повышается, чтобы обеспечить поглощение окружающих жидкостей.
Происходит преобразование микротубулярного аппарата цитоскелета. Ядерная мембрана растворяется
и путем перераспределения микротрубочек образуется веретено деления.
Метафаза II
В метафазе II хромосомы уже более упорядоченно расположены в области экватора, и два конца
веретена расположены на двух противоположных полюсах (как в метафазе I). Главным отличием от
метафазы I является образование двух кинетохор в сестринских хроматидах, которые в данном случае
повернуты по направлению к противоположным полюсам.
9. Анафаза II
В ходе происходящей в это время анафазы II две сестринские хроматиды (1) каждой хромосомы
разъединяются, как в митозе.
Расхождение начинается в кинетохорах (2) – точке прикрепления тянущих нитей центрального веретена.
Отсюда хромосомы медленно растягиваются по направлению к центриолям (4), расположенным на
полюсах клетки, двигаясь вдоль микротрубочек (3), которые создают эффект тяги за счет сокращения.
Микротрубочки (5), которые не соединены с хроматидами, теперь становятся длиннее, таким образом
увеличивая расстояние между центриолями и растягивая клетку. В области экватора становится видна
начинающаяся стадия образования борозды деления (6).
10. Телофаза II и цитокинез II
Расщепление и деление двух клеток, образовавшихся во время первого деления мейоза, теперь приводит
к образованию четырех гаплоидных клеток (1) с различными генетическими комбинациями, образовавшихся
в результате случайного распределения хромосом и кроссинговера. Это объясняет тот факт, что сибсы
(родные братья и сестры) не идентичны: один ребенок больше похож на отца, другой - на мать. Также
возможно проявление признаков прародителей.
46
Page 47

®
中文
减数分裂,或者叫做成熟分裂,是细胞分裂的一种特殊类型。根据生物体或者细胞类型的不同,减
数分裂需要的时间可能会是几天或者几年,减数分裂最终会导致性细胞的产生(配子)。通过减数
分裂,每个前体细胞会产生四个配子。一般而言,有两种类型的配子。大的,不动的细胞被称为卵
细胞或者卵母细胞,而小的,运动的细胞叫做精子或者精母细胞。
卵细胞是在卵巢中,由前体细胞经减数分裂形成。人类的卵细胞在胚胎的时候(妊娠3到8个月的时
候)已经开始发育成熟,但是,细胞会保持在减数分裂的中间相,直到个体达到性成熟。从那时
起,一些未成熟的卵细胞就会在激素的控制下,在规律的间期完成减数分裂。达到性成熟的时候,
人类精子就会在睾丸中有规律地开始变得成熟。在此情况下,一次完整的减数分裂大概是20-24天。
通常,体细胞(如配子的前体细胞)包含两倍的(二倍体)染色体组,其中的一半来源于母亲,而
另一半来源于父亲。
因此,染色体中会存在两个副本,就像一对同源染色体。
与之形成对比的,配子只包括一个单个的(单倍体)染色体组。换句话说,卵细胞与精子只是包括
父亲或者母亲的一半的遗传信息,因此,当两个细胞在一起的时候,就能够形成一个完整的带有二
倍体染色体组分的新细胞(受精卵)。
因此,减数分裂的目的就是为了将配子前体细胞正常的二倍体染色体组减少为单倍体的补体,从而
建立任何有性生殖的基础。减数分裂另一个更进一步的功能是为了将遗传信息混合。这是通过如下
两个机制完成:
1. 母亲和父亲的同源染色体随机分配到生成的生殖细胞中
2. 同源染色体之间基因的互换(基因重组)
(这两种机制的基本程序将在下面各个阶段中有描述)。
减数分裂
人类的单倍体中,有23对同源染色体,单个同源染色体的随机分配会造成2
变化的可能性。通过同源染色体之间的基因互换,变化的数量会进一步的升高。
在减数分裂之前,前体细胞是处于分裂间期的,分裂间期是指在两个细胞(有丝分裂或者减数分
裂)分裂之间的时期。
分裂间期包括3个阶段:
• G1相(合成前期)细胞生长的阶段。
• S相(合成期)在这一阶段,中心粒与DNA(去氧核糖核酸)已经开始复制。
• G2相(合成后期)这个阶段是从分裂相中分离出的DNA合成的末尾。而且,中心粒的复制也是在
这个阶段完成。
减数分裂,是在分裂间期后的一个阶段,包括两个连续的成熟分裂(减数分裂的),中间还被一个
短的、特殊的分裂间期所分割(分裂间期)。
与有丝分裂一样,在每一个减数分裂中,都分化为几个分裂阶段:
第一减数分裂:
• 前期I(4个小部分:细线期、偶线期、粗线期、带有终变期的双线期)。
• 中期I
• 后期I
• 末期I
• 胞质分裂I
分裂间期
第二减数分裂:
• 前期II
• 中期II
• 后期II
• 末期II
• 胞质分裂II
23
,即8.4x106种不同基因
47
Page 48

®
中文减数分裂
3BScientific®减数分裂模型系列(产品号R02)以及减数分裂的壁挂图(V2051M,V2051U)显示了典型
的哺乳动物细胞,放大倍数大约为10,000倍。在模型/图解的下三分之一,还有关于细胞器的描述。
3BScientific®减数分裂模型系列是放置在一个贮存系统之内的,有一个悬挂装置来装备。这样,为了
节省空间,模型系列可以很简单地悬挂在墙上。在模型的后部还有磁铁,因此,模型可以被安放在
教室里的磁性板上,以满足教学的要求。
在这份说明的末尾,您将会看到所包括的10个阶段的图解。您可以将这些图解复印来用于教学使
用。通过彩色的、带有标记的以及正确安排的各个的阶段,您的学生能够很容易地复习和记忆他们
所学习的。
在网上也可以找到各个阶段的免费彩图,网址是www.3bscientific.com。
1.分裂前期,G1相的阶段
在细胞内部,可以见到带有核仁(1)及其核膜(2)的细胞核。核仁内还包括尚未螺旋的带有遗传信息
的DNA(3)。
细胞自身通过一种极细的管,被称作微管(4),在细胞质内延伸来保持它的稳定性和形状。微管控制
细胞运动和细胞内的运输过程。
在细胞浆内,可以看到内质网(5),这是一个缠绕的管状系统,主要负责脂类合成、离子的存储和重
设计,以及运送某种蛋白。粗面内质网的膜上附着有核糖体,核糖体的功能是通过内质网来合成蛋
白。
高尔基复合体(或高尔基器)(6)也被称为“细胞腺体”,它是由层状的空泡层叠堆积组成(高尔基
囊泡),当小囊泡变小并且“干瘪”的时候(高尔基小泡)(7),它就会膨胀。高尔基复合体接受膜
成分和来自内质网的酶。它的主要功能是收集、包裹和运输分泌物和产生溶酶体(消化小泡)(8)。
溶酶体的主要工作是让细胞成分崩解(细胞内消化)。线粒体(9)负责生产细胞的能量。
中心粒(10)的工作是建成卵裂纺锤体,它是空的圆柱状,由许多纵形管状物(微管)组成。
2.前期I
第一减数分裂的前期是减数分裂中耗时最长的部分,在这个阶段中,染色体和染色质会改变它们的
结构,并且按照一个特殊的顺序排列在细胞核当中。因此,前期I可以分裂4个小部分(细线期、偶
线期、粗线期和带有终变期的双线期)。与有丝分裂前期大不相同的是,减数分裂前期I能够耗时几
天、几周、几个月或者几年,而有丝分裂只是持续几个小时。
细线期
在前期I的开始阶段(细线期),能够看到核仁(1)与核膜(2)。此时可以见到染色体(3)呈现单个的、
长的、细丝装。它的末端连接在核膜上,在分裂间期,每一个染色体都已经复制,它有两个姊妹染
色单体组成,但是它们彼此非常靠近,以至于很难被区分。
在分裂间期,中心粒也会被复制,两个中心粒(4)开始朝向两个相反的方向,即细胞的两极移动。
在它们中间开始构建中心纺锤体(5),中心纺锤体是由许多微管组成。
3.偶线期和粗线期
一对染色体中,一个母体的(1)和一个父体的(2)同系物(包括两个姊妹染色单体)采用不同的颜色
来表示,代表不同的同源染色体(整体2x23)。
偶线期
一旦同源染色体并排的排成队列形成联会丝复合物(3)(同源染色体配偶体平行排列),偶线期就开始
了。这一过程通常是以染色体的末尾开始,并继续下降至另一端,这与拉链很相似。染色体配对
(联会)是以非常高的精确度发生的,这是为了同源染色体的匹配基因能够直接地彼此面对。
这对于基因部分的重组交换(染色体交换)是非常重要的。
在减数分裂I期中,同源染色体的配对通常涉及二价染色体,但是,既然每个同源染色体都包括接近
的姊妹染色单体,它们就也有可能会涉及四倍体。
粗线期
一旦所有的联会复合体都得到充分地发育,换句话说,同源染色体都已经排好队列,粗线期就开始
48
Page 49

®
中文 减数分裂
了。自此以后,在联会复合体的间隔处,重组结(4)变得可见,在此处开始基因片段的交换。
4.双线期
在一些基因片段已经交换之后,同源染色体(1)越来越分离,只是在一个或者多个交换点的地方保持
连接(染色体交叉点或交叉)(2)。染色体交叉点是早期进行基因重组的地方(父亲的和母亲的遗传信
息交换的地方)。卵细胞能够保持在双线期几个月,甚至几年。
5.终变期
当染色体从核膜(1)中脱离的时候,减数分裂的前期I的终变期就开始了。染色体凝集,具有着丝粒
(短DNA序列,具有高的AT水平)(2)的姊妹染色单体变得可见。在基因片段交换的地方,非姊妹染
色单体通过染色体交叉点(3)保持连接。前期I随后的阶段是中期I,保持在这个点的减数分裂阶段,
不到完成整个减数分裂时间的10%。
6.中期I
在从前期I进入到中期I的过渡期间,成对的中心粒(1)已经到达细胞相反的两极,纺锤体已经形成,
核膜(2)已经溶解。染色体在赤道水平列队,形成所谓赤道板。从顶部看,染色体成一个星形(单星
体或者“母星体”)。着丝点(3)是一个蛋白质复合体,它已经形成着丝粒。减数分裂的中期I的一
个特质就是,每个姊妹染色单体对上的着丝点看起来像是已经融合。中心纺锤体的微管(4),此时已
经将其自身精确地连接到每一对姊妹染色单体上(5),因此所有的点都在同一个方向。染色体交
叉点(6)还是完整的。它们在将同源染色体正确的排列在赤道水平这一过程中扮演者十分重要的角
色。
内质网(7)和高尔基复合体(8)现在几乎完全被溶解了。
7.分裂后期I
在减数分裂的后期I,姊妹染色单体中的同源染色体(1)分离,这与有丝分裂中的不同。
在这一过程中,染色体的交叉点已溶解,在此之前,是它将母体和父体的同源染色体固定在一起
的。一些突变的有机体,有丝分裂染色体交换只是发生在有限的水平,会出现没有染色体交叉点的
染色体对。这些染色体对通常没有充分分离(对偶染色体不分离),从而导致子细胞存在太多或者
太少的染色体。此种畸形被称为染色体数字畸变,染色体数字畸变可能会引起畸形。
间断是在着丝点开始(2),着丝点是中心纺锤体收缩纤维连接的地方。从这里,染色体慢慢朝向
位于细胞两极的中心粒(4)牵拉沿微管(3)移动,当微管变短的时候,就会产生牵拉的作用。没有
连接到染色体的微管(5)现在变得更长了,因此增加了中心粒和正在延伸的细胞之间的距离。在
赤道水平上,卵裂沟的开始阶段(6)变得可见。在前期期间的交叉过程以及父体和母体染色体到
细胞两极的随机分布,导致了遗传信息的变化(请见介绍部分)。
8.末期I,胞质分裂I,分裂间期,前期II和中期II
末期I和胞质分裂
在末期I,纺锤体的裂解和一个环形的狭窄带(1)在赤道水平形成。此外,形成了一层很薄的核
膜(2)。在随后的胞质分裂阶段,细胞体恰好在中部分裂,即两个新的子核(3)之间的环状狭
窄处。每个子核都包括母体和父体的染色体组,这些组分通过交叉的过程已经有了轻微的改变,
DNA已经在复制中呈现,即包括两个姊妹染色单体(4)的一个染色体。
内质网(5)和高尔基复合体(6)都已经返回到它们原来的形状和大小。
在胞质分裂的末期,完成了第一减数分裂。
49
Page 50

®
中文减数分裂
减数分裂间期
第一和第二减数分裂是通过一个短暂的休眠期来分开(分裂间期)。但是,不包括两个染色单体的
染色体的复制(无S阶段)。每个染色体的两个姊妹染色单体都会通过着丝粒(7)来保持连接。
减数分裂II
第二减数分裂类似于有丝分裂(通常的核及细胞的分裂)。它因此也被叫作均等分裂。既然染色体
在前面的分裂间期中没有被再次复制,那么随后的第二减数分裂将包括的基因信息减少到了单倍染
色体组。
前期II
前期II是与有丝分裂最相似的一个阶段,在所有的生物体内发生的都非常快。细胞表面的通透性增
加,允许周围的液体吸收。细胞骨架的微管装置被重新组织。核膜溶解,由重排的微管构建成纺锤
体。
中期II
在中期II,染色体被再一次的重排于赤道水平,纺锤体的两端也定位于相反的两极(像在中期I中一
样)。此阶段与中期I的主要差别在于在姊妹染色单体形成两个着丝点,并朝向相反的极向。
9.后期II
在后期II,如同有丝分裂一样,每个染色体的两个姊妹染色单体(1)会分离。
分离是从着丝点(2)开始,着丝点是中心纺锤体的牵拉纤维附着的地方。从这里,染色体被慢慢的朝
向中心粒(4)牵拉,沿着微管(3)向前移动,而中心粒坐落于细胞的两极,当微管变短的时候,就会
产生出一个牵引力。不与染色体连接的微管(5)则变得更长了,因此,就增加了中心粒与正在延
伸的细胞之间的距离。在赤道水平,卵裂沟(6)的开始阶段变得很明显。
10.末期II与胞质分裂II
在第一减数分裂期间,两个细胞的卵裂与分割导致了四个单倍体细胞(1)的产生,这四个单倍体带有
不同的遗传组成,这是由于随机染色体分配与交叉所导致。这就能够解释,为什么同胞并不是一样
的:一个孩子有更多来自父亲的特征,而另一个孩子有更多来自母亲的特征。祖先的特征重现也是
有可能的。
50
Page 51

51525354555657585960616263
Page 52

Page 53

Page 54

Page 55

Page 56

Page 57

Page 58

Page 59

Page 60

Page 61

Page 62

Page 63

Page 64

R02/V2051_1000522/1001210-03/12-3
®
3B S C I E NT I F I C
3B Scientific GmbH
Rudorffweg 8 • 21031 Hamburg • Germany
Tel.: + 49-40-73966-0 • Fax: + 49-40-73966-100
www.3bscientific.com • 3b@3bscientific.com
© Copyright 2009 / 2012 for instruction manual and design of product:
3B Scientific GmbH, Germany
P R O D U C TS
5002142
 Loading...
Loading...